Книга: Происхождение языка: Факты, исследования, гипотезы
Назад: Глава 4 Коммуникация в мире животных
Дальше: Глава 6 Гипотезы о происхождении языка
Глава 5
Как получить новое, унаследовав старое?
Человекообразные обезьяны в языковых проектах демонстрируют удивительные способности, но все же овладеть языком на уровне взрослого человека оказываются не в состоянии. Не демонстрируют они сравнимых с человеческими языковых способностей и в природе. Это значит, что наши общие с обезьянами предки еще не имели языка. Откуда же взялся язык, если его раньше не было? Человек, как известно, «произошел от обезьяны». Значит, человеческий язык каким-то образом получился в результате эволюции из коммуникативной системы наших «обезьяньих» предков.
Еще до недавних пор многие специалисты представляли себе это эволюционное развитие примерно так: у кого-то из предков человека появилась генетическая мутация, в результате которой он обрел языковую способность. На это как будто бы указывают данные специфического нарушения речи (англ. SLI — specific language impairment), обусловленные повреждением гена FOXP2 на седьмой хромосоме. Люди, страдающие этим наследственным заболеванием, испытывают трудности с грамматикой своего родного языка — примерно такие же, которые многие из обычных людей испытывают с грамматикой иностранного. Они не чувствуют некоторых различий между регулярной и нерегулярной морфологией, иногда оказываются не в состоянии построить по общему правилу форму от незнакомого слова. С. Пинкер, опираясь на исследования Мирны Гопник, приводит случай, когда женщина, страдающая SLI, не справилась с так называемым ваг-тестом: человеку показывают изображение некоторого вымышленного существа и говорят, что оно называется ваг (англ. wug), а потом показывают картинку с двумя такими существами и просят закончить фразу Здесь два…. Для обычного четырехлетнего носителя английского языка не составляет никакого труда сказать, что на картинке изображены two wugs, но человеку, страдающему SLI, нелегко сообразить, какое правило надо применить в данном случае. Как показывают исследования Татьяны Владимировны Черниговской и Киры Гор, люди, страдающие SLI, как бы хранят формы слов в памяти в целом виде, почти не проводя их морфологического анализа. Возможно, похожим образом обстоит дело и с предложениями: понимание конструкций с необычным порядком слов вызывает у таких больных значительные трудности, «грамматические процедуры как бы симулируются за счет памяти и эксплицитных правил». Нарушается при SLI и фонетический уровень: как пишет Черниговская, у таких людей отмечаются «монотонная речь, нарушение речевого ритма и неправильное членение звукового потока; нарушение просодики, ударения как в частотных, так и в незнакомых, но ясных с точки зрения правил данного языка, словах; сложности произнесения многосложных слов (сокращение их за счет пропуска слогов)». Кроме того, у них несколько нарушен моторный контроль за мышцами рта — им сложно, например, высунуть язык или дунуть по команде.
Как показывают исследования, ген FOXP2 подвергся мутации в ходе эволюции человека: две из трех замен, происшедших в этом гене со времени существования общего предка человека и мыши, произошли после разделения гоминид и человекообразных обезьян.
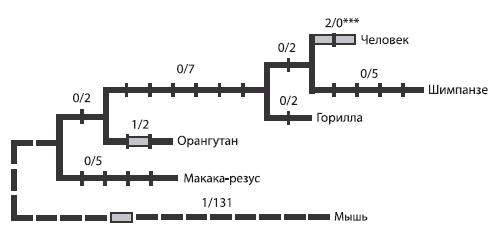
Рис. 5.1. Эволюция гена FOXP2. Цифра до знака дроби обозначает количество значимых нуклеотидных замен, после знака дроби — число незначимых («синонимичных»), то есть не приводящих к синтезу другой аминокислоты замен.
Не означает ли это, что причиной возникновения человеческого языка послужила одна-единственная макромутация, сформировавшая речевой аппарат, изменившая форму черепа, перестроившая мозг и «вложившая» в него языковую способность, как утверждали авторы некоторых работ? Так, по мнению Д. Бикертона, такая мутация вполне могла бы произойти у «митохондриальной Евы». А Тимоти Кроу, напротив, связывает языковую мутацию с Y-хромосомой, наследуемой исключительно по мужской линии.
Все же, по-видимому, идея о том, что человеческий язык мог возникнуть в результате единственной мутации, неверна (показательно, что в своей более поздней работе Д. Бикертон от нее отказывается). В гл. 2 мы видели, сколько различных механизмов — как анатомо-физиологических, так и когнитивных — должны слаженно работать, чтобы люди могли пользоваться языком. Органы, задействованные в обеспечении функционирования языка, возникают в процессе развития из разных эмбриональных зачатков — например, в развитии слухового анализатора (весьма ценного для звучащей речи) участвуют слуховая плакода, зачаток рудиментарной жаберной щели, элементы подъязычной и челюстной висцеральных дуг, мезодерма головы и невральная эктодерма. И это лишь один из множества необходимых для работы языка компонентов. Как пишет биолог-эволюционист Николай Николаевич Иорданский, «совершенно ясно, что невозможно возникновение всей этой сложной, но целостной системы как путем случайного подбора различных мутаций (например, при дрейфе генов), так и посредством одной крупной „системной“ мутации». Очевидно, что перестройка речевого аппарата и формирование нужных мозговых структур и всех прочих, столь многочисленных, сложно устроенных и хорошо сбалансированных приспособлений в результате случайной замены нуклеотидов в молекуле ДНК не более вероятны, чем сборка самолета смерчем, пронесшимся по свалке.
На это иногда возражают, что генетические изменения имеют так называемый «плейотропный» эффект — мутация, происшедшая в одном гене, может приводить к множественным последствиям в самых разных структурах организма. Но приводимые в литературе примеры плейотропного эффекта — это примеры разрушений. И действительно, нарушить работу сложного механизма, заменив в нем даже всего одну деталь на негодную, тем легче, чем сложнее механизм, но получить слаженно работающую систему, добавив «лишнюю» деталь, гораздо труднее — это понимает всякий, кто пытался если не переделать собственными руками велосипед в мотоцикл, то хотя бы установить на свой компьютер программу, написанную для другой операционной системы.

Рис. 5.2. Разрушить такое сооружение, вытащив любую карту из нижнего яруса, очень легко, а вот создать нечто подобное, подложив карту в нижнюю часть кучи карт, невозможно.
Муха-дрозофила, получившая в результате мутации лапку на месте усика-антенны или четыре крыла вместо обычных двух (см. фото на вклейке), безусловно, является другим существом, сильно отличающимся от своих родителей (в последнем случае это различие на уровне отрядов). Но никакой выгоды для мухи в этом нет: весь ее организм рассчитан на два крыла, пару жужжалец, две антенны и шесть лапок на своих местах. Такого рода мутации не создают ни систем управления, которые бы позволяли «лишним деталям» слаженно взаимодействовать с прочими частями организма, ни поведенческих программ, которые бы обеспечивали эффективное использование новообретенных признаков.
Против идеи единственной макромутации говорит и то, что генов, так или иначе принимающих участие в обеспечении функционирования языка и подвергшихся изменениям в ходе эволюции человека, обнаруживается с каждым годом все больше. Таков, например, ген, кодирующий альфа-текторин (один из важных компонентов текториальной мембраны внутреннего уха). В той же статье в журнале «Science», написанной большой группой авторов, перечисляются и другие гены, претерпевшие изменения в ходе развития человеческого слухового анализатора, — DIAPH1, FOXI1, EYA4, EYA1 и OTOR. Явно служил мишенью отбора в гоминидной линии ген HAR1 (точнее, это не ген, а участок гена HAR1F, кодирующего регуляторную РНК), — человек и шимпанзе имеют в нем 118 отличий, тогда как шимпанзе и курица — всего два. Ген FOXB1 необходим для обеспечения оперативной памяти — а при сбоях в оперативной памяти человек не сможет полноценно пользоваться языком (например, он не сможет обобщить синтаксические правила, определенные на достаточно длинных предложениях). Определенную роль в овладении языком играет ген c-fos, который включается в мозге в ответ на новизну (т.е. в том случае, когда поступающие в организм внешние стимулы не находят соответствия в том, что уже хранится в памяти). Важны для языка и гены MCPH1 и ASPM, регулирующие объем мозга, и многие другие гены — все те, которые принимают участие в обеспечении механизмов памяти, планирования поведенческих программ, генерализации правил, формировании в ходе развития организма (в том числе и после рождения) необходимых анатомических структур и т.д. Можно, по-видимому, утверждать, что язык, как и многие поведенческие признаки, имеет полигенное наследование.
Кроме того, гены в принципе не могут кодировать грамматические правила — каждый ген является единицей транскрипции, отрезком ДНК, на базе которого синтезируется одна молекула РНК (информационная, рибосомная, транспортная или регуляторная). Записать на него программу типа «ставь подлежащее перед сказуемым» или «ставь подлежащее после сказуемого» (с тем чтобы потом можно было «перещелкнуть переключатель» между этими двумя возможными вариантами), по-видимому, технически невозможно.
Соответственно, присущая человеку «врожденная языковая способность» предстает не как набор предзаданных правил Универсальной Грамматики, а как стремление искать структуру и «заполнять пустые клетки» в логически — по презумпции — организованной «таблице» коммуникативной системы. Авторы многих работ признают, что врожденными являются не свойства грамматики языка, а свойства механизма его усвоения, не детали коммуникативной системы, а «механизмы их выведения из поведения окружающих». В книге Р. Джакендоффа предпринимается попытка совместить идею врожденности УГ с уже не подлежащим сомнению фактом отсутствия специального «языкового органа» (который должен был бы, согласно гипотезе Н. Хомского, ее содержать). Универсальная Грамматика в интерпретации Р. Джакендоффа не является «монолитной способностью», а состоит из нескольких автономных слоев, представляя собой не набор правил, а «устройство для овладения языком» (англ. LAD — Language Acquisition Device). При этом наследуется УГ тем же способом, что и поведение.
Наличие универсальных закономерностей (как в области грамматики, так и в области «универсального словаря» — «мыслекода») объясняется, вероятно, тем фактом, что у всех людей много общих перцептивных, поведенческих, эмоциональных предрасположенностей, а даже слабые предрасположенности, если они есть всегда, ведут к значительным последствиям. Следует, впрочем, отметить, что универсальность этих закономерностей относительна: хотя они представлены во многих языках, но едва ли не для каждой из них можно найти такой язык, в котором она отсутствует.
Что же касается мутации в гене FOXP2, то она в любом случае произошла задолго до «митохондриальной Евы» — такой же, как у нас, вариант этого гена был обнаружен у неандертальца.
Вообще, следует отметить, что от гена и мутации в нем до внешнего (или, как говорят биологи, фенотипического) свойства — длинный путь. В клетке существуют специальные ферменты, которые исправляют ошибки, возникающие при копировании ДНК (так что в конечном счете остается примерно 1 ошибка на 10 млн. пар нуклеотидов). Многие из остающихся замен синонимичны: например, если в последовательности ЦЦТ заменить последний нуклеотид Т на А, Ц или Г, полученный триплет все равно будет кодировать ту же самую аминокислоту — глицин. Замена одной аминокислоты в белке может никак не повлиять на его функцию. Но даже в том случае, если мутация окажется значимой, другие гены, имеющиеся у организма, могут усилить или ослабить ее действие и даже полностью свести его на нет. Так, например, гиляцкие лайки — совершенно бесстрашные собаки, несмотря на то, что генетически они предрасположены к трусости. Их мало кому доводилось видеть испуганными, поскольку эти лайки очень маловозбудимы — их, как говорится, не проймешь. Генетические характеристики, обусловливающие малую возбудимость, блокируют проявление трусости. Таким образом, даже если предположить, что одна мутация вызвала у человека некую «предрасположенность» к языку, в отсутствие множества других изменений эта предрасположенность с большой вероятностью вовсе никак бы не проявилась.
У видов, использующих исключительно половое размножение, в каждом следующем поколении набор генов, которые могут влиять на внешнее проявление свойства, связанного с мутантным геном, будет изменяться, поскольку половина генетического материала достается организму от одного из родителей, а половина — от другого. И проявиться в виде фенотипического свойства может лишь та мутация, которой не помешает в этом вторая половина генома. Поэтому, например, у женщин — носительниц гемофилии внешних признаков этого заболевания не наблюдается: работа второй X-хромосомы, не затронутой мутацией, обеспечивает им в конечном итоге вполне удовлетворительный уровень свертываемости крови.
Усложняет связь между генами и фенотипическими характеристиками и тот факт, что новые свойства, отсутствовавшие у родителей, организм может получить не только в результате мутаций, но и в результате ненаследственных изменений — модификаций или морфозов. Например, попадание некоторых веществ в организм вполне генетически нормальной беременной женщины может повлечь развитие у плода врожденных уродств. Дрозофила с четырьмя крыльями вместо двух может появиться не только вследствие мутации (вернее, двух мутаций — bithorax и postbithorax), но и том в случае, если на ранней личиночной стадии она попадет под воздействие эфира или высокой температуры. Такое изменение не будет наследоваться, оно будет, как говорят биологи, фенокопией данной мутации (а сама мутация, соответственно, генокопией данного морфоза). Между внешними проявлениями мутаций и модификаций существует, как показал биолог-эволюционист Иван Иванович Шмальгаузен, глубокий параллелизм, основанный на том, что набор белков, которые может синтезировать клетка, далеко не безграничен: чаще всего речь идет о том, что характерные для организма белки начинают синтезироваться с другой интенсивностью или в другие сроки, и именно этим определяется набор потенциально возможных отклонений от исходного фенотипа.
Роль внешней среды для формирования тех или иных внешних признаков нередко оказывается весьма существенной даже для тех организмов, которые обладают значительной автономностью развития: существуют некоторые элементы среды, которые полезно «заметить» для повышения приспособленности, и естественный отбор благоприятствует тем организмам, в чьем развитии заложена программа реагировать на них определенным образом. Например, такое свойство, как способность к импринтингу, полезно, поскольку оно позволяет птенцу выработать эффективную реакцию на конкретную особь — собственного родителя. Более того, в индивидуальном развитии (онтогенезе) существуют специальные чувствительные периоды, когда развивающийся организм наиболее восприимчив к воздействиям определенного типа. Есть чувствительный период и в усвоении языка (см. гл. 1). Если человек не овладел языком в этот период, он, скорее всего, уже не сможет стать полноценным носителем; «вероятные причины этого — возрастные изменения в мозге, такие как уменьшение уровня метаболизма и количества нейронов на протяжении младшего школьного возраста и достижение наименьшего количества синапсов и уровня метаболизма примерно в пубертатном возрасте».
Внешние по отношению к организму факторы могут существенно изменить проявления того, что закодировано в его генах. Например, добиться «пассивно-оборонительной» (по терминологии специалиста по поведению животных Леонида Викторовича Крушинского), т.е. трусливой, реакции от гиляцких лаек можно, если искусственно поднять уровень их возбудимости путем введения кокаина. Под действием факторов внешней среды может изменяться в том числе устройство мозга — как уже говорилось в гл. 2, нейроны его коры могут перепрофилироваться. Если у младенца, еще не овладевшего языком, удалить все левое полушарие (такое иногда приходится делать по медицинским показаниям), он сможет впоследствии научиться говорить — языковые центры разовьются в сохранившемся правом.
Более того, различные мутации при определенном подборе разных условий могут вызывать одинаковые внешние проявления. При изучении поведения лабораторных мышей было выяснено, что, например, разницу в уровне исследовательской активности у мышей двух различных линий можно нивелировать (и даже заменить на противоположную) путем введения некоторых фармакологических веществ. Роберт Трайон вывел линии «умных» и «глупых» крыс — первые значительно быстрее, а вторые значительно медленнее обучаются находить пищу в 17-тупиковом лабиринте. Но в 16-тупиковом лабиринте крысы обеих линий демонстрируют одинаковые успехи.
Таким образом, то, каким в итоге окажется человек (в том числе в аспекте владения языком), зависит от генов его родителей и их конкретной комбинации (например, от родителя, имеющего генетическую предрасположенность к SLI, ребенку может передаться не дефектный, а второй, нормальный вариант гена FOXP2), от характеристик цитоплазмы яйцеклетки (так называемый «материнский эффект» в узком смысле), от условий внутриутробного развития (например, употребление матерью алкоголя может крайне негативно сказаться на развитии мозга плода, что может помешать ему впоследствии успешно овладеть языком) и, наконец, от тех воздействий, которые окажет на него окружающая среда после рождения (например, сильный испуг может сделать человека заикой). Именно поэтому, кстати, при клонировании невозможно получить точную копию родителя — для этого пришлось бы воспроизвести в точности не только гены, но и все релевантные условия, регулирующие индивидуальное развитие. И именно поэтому, сколь бы умны ни были родители человека, сколь бы хорошо они ни владели языком, это не дает стопроцентной гарантии, что их потомки будут умными и красноречивыми.
Разумеется, гены в наследовании играют очень важную роль. Во-первых, они задают возможный спектр изменчивости: какие свойства можно получить в результате мутаций, а какие — нельзя. Например, невозможно получить синие глаза у дрозофилы (по крайней мере, среди изученных примерно двух миллиардов дрозофил синеглазые особи не попались ни разу). Не существует мутаций, которые бы случайным образом наделяли любые организмы любыми свойствами так, чтобы из них потом можно было отбирать оптимальные, — и в этом ответ на знаменитый вопрос «Почему у свиней нет крыльев?». Их нет не потому, что естественный отбор отбраковал всех крылатых свиней, появившихся в результате мутации, а просто потому, что набор генов, характерных для свиней (как и для всех других копытных), не предусматривает возможности появления крыльев — ни в качестве дополнительной пары конечностей, ни в качестве модификации уже имеющихся.
Во-вторых, гены определяют, на какие факторы внешней среды организм (в том числе в своем развитии) будет реагировать в большей, а на какие — в меньшей (в частности, в нулевой) степени и каков будет характер этой реакции. В-третьих, под генетическим контролем находится то, в какой период развития тот или иной фактор внешней среды будет оказывать сильное (подчас определяющее) воздействие на организм, а в какой — слабое (или вообще никакого). Общий генофонд популяции задает частоту, с которой будут возникать те или иные характеристики особей (как анатомо-физиологические, так и поведенческие), и даже частоту появления различных мутаций. В число факторов среды, оказывающих влияние на развитие организма на любой отдельно взятой стадии, входят результаты работы генов на стадии предшествующей.
Еще один существенный для понимания механизмов наследования момент состоит в том, что работа генов не заканчивается с рождением организма, и внешние условия могут оказывать на нее влияние. Например, как было выяснено недавно, не только гены могут управлять поведением, но и, наоборот, поведение может управлять генами. Американские исследователи Джин Робинсон, Рассел Ферналд и Дэвид Клейтон задались вопросом о механизме наследования родительской заботы у крыс. Оказалось, что этот феномен определяется не родительскими генами и не научением. Просто, когда крыса-мать облизывает своих детенышей, у них происходит метилирование определенных участков ДНК, что вызывает «выключение» ряда генов, и в результате изменяется реакция нейронов на некоторые гормоны, а это, в свою очередь, приводит в дальнейшем к увеличению уровня заботы выросших крысят о своих собственных детенышах. Метилирование ДНК — не генетический (а так называемый «эпигенетический») процесс, но при нормальном ходе событий (крыса-мать вылизывает своих детенышей, из них вырастают заботливые матери, которые, в свою очередь, вылизывают своих детенышей и т.д.) это обеспечивает вполне эффективное наследование данного поведенческого признака.

Рис. 5.3. Чтобы дети выросли заботливыми, их надо как следует вылизывать.
Все это показывает, что попытки однозначно определить, что же все-таки сыграло ключевую роль в возникновении языка — гены или среда (проблема, получившая в англоязычной литературе название «nature— nurture», «природа — воспитание»), едва ли могут привести к осмысленному результату. Скорее, на мой взгляд, разумно изучать взаимодействие генов и среды в этом процессе.
Эволюция живых организмов происходит путем естественного отбора. Но естественный отбор не работает с мутациями напрямую — ему доступны лишь фенотипы. Так, если бы дрозофилам по какой-то причине стало выгодно иметь четыре крыла, в выигрыше оказались бы не только носители соответствующего комплекса мутаций, но и те, кто получил дополнительную пару крыльев в результате морфоза (хотя, разумеется, последние не смогли бы передать эту особенность своим потомкам).
Новые фенотипические характеристики могут возникнуть не только непосредственно в результате мутаций, но и вследствие комбинационной изменчивости — гены родителей, работая вместе в одном организме, могут наделять его тем или иным свойством, которого не было в фенотипе ни у одного из родителей. Например, как показал Л.В. Крушинский, если скрестить бесстрашную немецкую овчарку с не менее бесстрашной гиляцкой лайкой, получится очень трусливая собака, поскольку щенок унаследует от родителя-лайки склонность к пассивно-оборонительной реакции, а от родителя-овчарки — высокую возбудимость. В каждой популяции циркулирует определенное количество вариантов (аллелей) различных генов, и признаки, получаемые в результате их комбинаций, хотя и не наследуются напрямую, тем не менее возникают у членов этой популяции вновь и вновь. Частота появления особей с теми или иными фенотипическими особенностями входит в набор характеристик вида (и может меняться со временем).
Для успешного прохождения отбора не имеет значения не только то, каковы в точности у особи гены, но и то, каковы ее отдельные свойства, — необходимо лишь, чтобы весь комплекс присущих особи индивидуальных признаков в комбинации с характеристиками той среды, где она живет, обеспечивал наилучшие показатели выживания и размножения. Например, в районах, где распространена малярия, у многих людей в генах заложена устойчивость к ней. Как выяснили генетики, обеспечивается она несколькими различными мутациями (а значит, формировалась в разных регионах независимо). При этом сама по себе особенность, кодируемая соответствующими генами, вредна — человек, получивший мутантные гены от обоих родителей, быстро умирает от серповидно-клеточной анемии. Однако в условиях значительной угрозы малярии плюс возможности комбинации с немутантными генами второго родителя такая мутация поддерживается естественным отбором. (Заметим в скобках, что отбор в данном случае закрепляет не только саму мутацию, но и уровень ее распространения — ненулевой, но и не стопроцентный.)
Любое отдельное свойство может быть в разных условиях и положительным, и отрицательным, и нейтральным. Например, густой волосяной покров для млекопитающих, живущих в холодном климате, является благом, поскольку защищает их от потери тепла, но для земноводных с их кожным дыханием или, скажем, для бегемотов, которым крайне важно избегать перегрева, был бы смертелен. Склонность издавать громкие звуки при виде опасности повышает выживаемость у общественных животных, но понизила бы ее у животных, ведущих одиночный образ жизни.
Поэтому при возникновении новых фенотипических особенностей — как вследствие мутаций, так и в результате модификаций и комбинирования родительских генов — лучше приспособленным оказывается не тот организм, у которого соответствующие показатели выражены в наиболее сильной степени, а тот, который смог наиболее успешно «вписать» их в контекст как своих собственных характеристик, так и условий окружающей среды.
У каждого вида есть так называемая «норма реакции» — это, по определению теоретика эволюции Алексея Сергеевича Северцова, «пределы, в которых может изменяться фенотип без изменения генотипа»; в этих пределах организмы могут приспосабливаться к условиям среды. Когда какие-то фенотипические черты (возникающие в результате комбинации родительских генов, или реакции на какие-то факторы внешней среды, или и того, и другого), а точнее комплексы черт, в определенных условиях оказываются полезными, организмы, их имеющие, лучше выживают и размножаются, и отбор благоприятствует тем, в чьем потомстве эти свойства проявляются с большей вероятностью (и большей надежностью). Соответственно, рано или поздно вырабатывается механизм, с достаточной надежностью обеспечивающий совместное наследование удачно сочетающихся между собой признаков — возникают мутации, повышающие вероятность передачи следующему поколению всего этого комплекса целиком (в пределе — до единицы), и эти мутации закрепляются отбором.
Подобные мутации могут происходить в генах, которые регулируют активность других генов. И, вероятно, не случайно, что многие генетические отличия человека от шимпанзе связаны именно с имеющейся в геноме системой регуляции. Так, например, американский генетик Грегори Рэй и его коллеги недавно обнаружили, что у нас имеется мутация в регуляторной области гена, кодирующего белок продинорфин. Из этого белка в организме образуются эндорфины — вещества, участвующие в регуляции эмоций, влияющих на социальное поведение, формирование привязанностей, способности к обучению. Сам белок продинорфин у человека и шимпанзе идентичен, но у человека его синтезируется несколько больше (примерно на 20%), а кроме того, по-видимому, различаются реакции на внешние стимулы — в каких случаях продинорфина будет синтезироваться больше (и насколько), а в каких — меньше. А это, как пишет А.В. Марков, «могло серьезно изменить мотивацию человеческих поступков, наши желания и „жизненные цели“». Группа молекулярных биологов из США и Австралии, сравнив уровень активности 907 генов у человека и нескольких видов обезьян, обнаружила, что многие гены в ходе эволюции разных групп приматов увеличили или уменьшили свою активность. При этом среди генов, активность которых в человеческой линии повысилась, оказалось много таких, которые участвуют в регуляции активности других генов. Регулятором (причем регулятором высокого уровня — т.е. таким, который регулирует активность других генов-регуляторов) оказался и знаменитый ген FOXP2. Начинающийся с него «генетический каскад отвечает за нормальное развитие мозга, в особенности тех отделов, от которых зависит координация движений и, в том числе, артикуляция речи».
В любой популяции за счет комбинирования генов (обеспечиваемого половым размножением) и воздействия окружающей среды появляются различные фенотипы с различной частотностью. Если какой-то фенотип (именно фенотип в целом, обладающий некоторым балансом свойств) оказывается очень удачным, то преимущество получит та популяция, где такой фенотип (вне зависимости от его генетической основы) будет появляться чаще. И если перемещение и скрещивание особей внутри этой популяции происходит существенно чаще, чем между этой популяцией и соседними, то популяция, где чаще появляется удачный фенотип, будет производить большее количество потомства и, соответственно, «экспортировать» особей в другие популяции. Даже если среди этих особей будет не очень много носителей удачного фенотипа, принесенный ими генетический «багаж» будет способствовать тому, чтобы удачный фенотип и на новом месте появлялся с большей частотностью. И таким образом генофонд, способствующий более частому появлению удачного фенотипа, будет распространяться.
С точки зрения стороннего наблюдателя эволюция нередко выглядит как своего рода баланс спроса и предложения: если есть на что-то спрос, то будет и предложение, если формируются условия, в которых тот или иной признак мог бы оказаться полезным для выживания и размножения, он с большой вероятностью у кого-нибудь появляется. Например, если возникает новый вид насекомых, тут же возникает и паразит, специализированный именно к данному виду, если на какой-то территории появляется новое цветковое растение, непременно находится тот, кто начинает его опылять, им питаться и т.д. Когда люди стали осваивать животноводство, у них быстро распространились мутации, дающие возможность переваривать свежее молоко во взрослом возрасте — одна из них возникла в Европе, другая — на Ближнем Востоке, третья — в Африке. Подобные мутации конечно же иногда возникали (и по сей день возникают) и в других человеческих популяциях, но при отсутствии молочного животноводства они не дают никакой выгоды, поэтому они не закрепились естественным отбором.
Разумеется, новое свойство, полезное в возникших условиях, может и не появиться, — но тогда данный вид просто не сможет освоить соответствующую среду обитания, тип пищи или т.п. — и мы никогда не узнаем, что этот признак мог бы оказаться выгоден данному виду.
Соответственно, гипотеза о происхождении языка должна уметь отвечать на вопрос о том, каков же был тот «спрос» окружающей среды, на который гоминиды ответили таким «предложением». Язык — достаточно большое по объему, сложно устроенное и энергоемкое, а с точки зрения некоторых анатомических особенностей даже просто вредное образование. Если естественный отбор не отбраковал такое «излишество», значит, оно дает какие-то эволюционные преимущества. И современные авторы гипотез о происхождении языка стремятся установить, чем язык мог быть полезен (в некоторых работах это даже оговаривается специально).
Какие же эволюционные преимущества дает человеку язык? На этот счет было высказано немало гипотез. Джон Кребс и Ричард Докинз полагают, что язык развился как средство манипулирования сородичами (эта гипотеза и поныне имеет сторонников). Действительно, иногда стать объектом манипуляции выгодно: например, если цель манипулятора — добиться успеха на охоте за счет координации совместных действий. Но все же, как кажется, это не могло быть основным фактором глоттогенеза: в силу того, что объект манипуляции не осознает воздействия, он будет в равной мере склонен выполнять как обоюдно выгодные действия, так и действия, приносящие пользу лишь манипулятору. В таких условиях подающий сигналы будет стремиться к тому, чтобы добиваться от сородичей максимального количества действий, выгодных только ему самому. А тогда, как справедливо замечает Пинкер, преимущество получили бы те, кто не владеет языком и тем самым недоступен для манипуляций. У Т. Дикона развитие языка предстает как гонка вооружений: особи стремятся всё более эффективно обманывать других (к собственной выгоде) и всё более эффективно распознавать обман сородичей (чтобы самим не быть обманутыми). Связь языка с обманом прослеживается и у Д. Бикертона: согласно его гипотезе, большую роль в формировании языка сыграли «гены детектора лжи». Но и к этим идеям замечание Пинкера вполне применимо: особей, не владеющих языком, обмануть при помощи языковых средств невозможно. По мнению Джеффри Миллера, язык развился в первую очередь как средство продемонстрировать большие умственные способности при половом отборе. Но время, когда начинает действовать половой отбор, достаточно далеко отстоит в жизни человека от периода овладения языком, а двух-трехлетний ребенок вряд ли станет заботиться о том, что понадобится ему лишь через многие годы. Для устранения этого противоречия Дж. Локк выдвинул гипотезу, что более ранние стадии овладения языком подстроились под то, чтобы ко времени действия полового отбора все необходимые характеристики языка были в наличии. Близка к этому идея Жана-Луи Десаля о том, что главным назначением формировавшегося человеческого языка было повышение статуса говорящего: превосходя других в особом «спорте» — разговоре — сообщением релевантной и правдивой информации, человек обеспечивал себе более высокое место в иерархии, а значит, лучший доступ к ресурсам, половым партнерам, коалициям особей, которые встанут в данном сообществе у власти.
Действительно, этнографами зафиксирована практика «словесных поединков», да и в нашем нынешнем обществе (как вполне справедливо отмечает Десаль) человек пользуется бóльшим или меньшим уважением в зависимости от качества, уместности, интересности своих реплик, остроумия, проявляемого в разговоре, и т.п.. Можно предположить, что речь в данном случае рассматривается как индикатор общих умственных способностей, которые недоступны непосредственному наблюдению: чтобы их оценить, надо видеть действия индивида, их результаты, а также иметь возможность сравнить, каков был бы результат других действий в соответствующих обстоятельствах. Но ситуации, в которые попадает человек, настолько многообразны, что найти в точности такую же ситуацию, чтобы проверить эффективность других действий, чаще всего оказывается невозможно, поэтому средство, позволяющее судить об умственных способностях индивида без непосредственной проверки, очень ценно. Таким средством оказывается речь — у кого она гибче, богаче, кто находчивее в словесном поединке, тот, весьма вероятно, имеет достаточно большое количество нейронных связей в мозгу и лучше умеет ими пользоваться, сопрягая разные элементы своего жизненного опыта, подчас даже необычным образом. Соответственно, резонно ожидать, что, столкнувшись с какими-то нетривиальными проблемами в реальной жизни, он сможет найти решение с той же легкостью, с какой подбирает нужные слова. Но в таком случае язык не мог возникнуть вне связи с общим развитием интеллекта, исключительно как средство повышения собственного статуса путем ведения беседы, поскольку индикатор, который врет, — вещь бесполезная (и даже вредная). Если бы язык развивался исключительно для демонстрации самого себя, его наивысшим воплощением был бы синдром Вильямса — сочетание болтливости, способности к беглой (и грамматически правильной) речи с глубокой умственной отсталостью. Против гипотезы о половом отборе как о главном факторе глоттогенеза говорит и то, что язык в равной мере развивается и у мужчин, и у женщин, тогда как для признаков-«излишеств», возникших в результате полового отбора, — таких, как приводимый обычно в пример павлиний хвост, характерна привязка к одному из полов: у пав-самок роскошного хвоста нет. У человека же, напротив, болтливость ассоциируется в большей степени с женщинами — мужчины во многих культурах ценятся не за слова (которых должно быть мало), а за дела. Признаки, сформировавшиеся для ухаживания, демонстрируются обычно противоположному полу (а своему — только по ошибке), люди же в высшей степени склонны беседовать с представителями пола собственного. Кроме того, для признака, сформированного исключительно половым отбором, странно выглядит неприятное свойство давать сбои в самый важный для ухаживания момент, а язык именно таков: при виде наиболее желанного представителя (или представительницы) противоположного пола у многих людей начинаются серьезные трудности с речью. Как писала древнегреческая поэтесса Сафо, «Лишь тебя увижу, уж я не в силах вымолвить слóва».
Все эти (и подобные им) гипотезы исходят из той общей идеи, что язык должен давать адаптивные преимущества единичной особи, прежде всего — особи, подающей сигнал. Как отмечают антропологи М.Л. Бутовская и Л.А. Файнберг, в таких работах «дарвиновская идея борьбы за существование… представлена лишь как бесконечная конкуренция между особями, направленная на извлечение максимальной выгоды для себя». Такие концепции нередко связываются с упрощенным пониманием сформулированной в свое время Ричардом Докинзом теории «эгоистичного гена»: единицей отбора является ген, и эволюционный выигрыш могут принести только те свойства, которые дают этому гену возможность оставить максимум своих копий — если нужно, то в ущерб всему остальному миру. Собственно, максимум копий — это и есть эволюционный выигрыш.
Но сторонники таких представлений нередко забывают о том, что даже самый «эгоистичный» ген копирует себя не в одиночку, а в «компании» других генов — своих соседей по геному. И эти соседи могут способствовать копированию «эгоистичного гена», а могут и препятствовать. Если «эгоистичный ген» принадлежит виду, размножающемуся половым путем, то в следующем цикле репродукции соседи будут уже другими. В таких обстоятельствах наиболее выгодная для «эгоистичного гена» стратегия — правильно выбирать соседей. А это значит, что он должен либо сам диктовать организму выбор оптимального полового партнера (то есть такого, чьи гены как минимум не помешают «эгоистичному гену» успешно копироваться), либо пристраиваться к другому гену (или комплексу генов), который умеет подавать организму нужные «эгоистичному гену» команды. Как показали эксперименты на дрозофилах, потомки особей, имевших возможность выбирать полового партнера, живут статистически значимо дольше, чем потомки особей, такой возможности лишенных.
Отметим еще раз, что оптимальным половым партнером будет не тот, у кого то или иное (пусть даже самое выгодное) свойство выражено в наиболее сильной степени, а тот, чьи гены сочетаются с генами данной особи наиболее удачным образом — дают возможность с максимальной вероятностью «произвести многочисленное, жизнеспособное и во всех отношениях успешное потомство при скрещивании не с кем-нибудь, а именно с „выбирающей“ особью» (то есть, например, немецкой овчарке не стоит выбирать в супруги самую бесстрашную гиляцкую лайку). Механизмы выбора оптимального полового партнера еще не до конца изучены, но есть данные, что значительную роль в этом играет способность организмов отличать «своих» от «чужих», тесно связанная с определенными компонентами иммунной системы. Как показали Александр Владимирович Марков и Алексей Михайлович Куликов, оптимальным (и, соответственно, наиболее привлекательным) половым партнером является та особь, которая — с точки зрения иммунной системы — отличается от данной не слишком слабо, но и не слишком сильно. При этом различные факторы внешней среды могут смещать оптимум в сторону большего или меньшего сходства. Например, высокая степень нестабильности окружающей среды благоприятствует выбору максимально генетически далеких половых партнеров: это повышает разнообразие доступных виду фенотипов, а значит, и вероятность, что при любых вариантах условий найдутся особи, которые сумеют в них выжить и дать потомство.
Но для того, чтобы можно было правильно выбрать полового партнера, необходимо не только уметь осуществлять выбор, нужно еще, чтобы вокруг циркулировало достаточное количество особей с похожими геномами, из которых можно было бы выбирать. А значит, эти особи должны, во-первых, жить достаточно близко друг от друга (или, по крайней мере, уметь собираться для размножения в нужное время в нужном месте), а во-вторых, при этом не очень мешать друг другу в борьбе за существование. Таким образом эгоистические интересы «эгоистичного гена» требуют поддержания устойчивого существования как минимум одной полноценной популяции. При этом популяция не должна состоять исключительно из близких родственников — более того, чем нестабильнее условия окружающей среды, тем сильнее увеличивается спрос на возможность мирного сосуществования неродственных особей. А значит, отбор будет поощрять гены, позволяющие минимизировать ущерб от соседствования с особями своего вида, а возможно, даже получить от этого ту или иную дополнительную выгоду. Наиболее простой способ достижения этого — родственный отбор: помощь родственнику ведет к тому, что гены, заставляющие помогать родственникам, лучше распространяются. Он описывается так называемым «правилом Гамильтона», выражаемым формулой
nrB > C,
где r — степень генетического родства того, кто помогает, и того, кому помогают, B — преимущество в борьбе за выживание или в производстве потомства, полученное адресатом альтруистического акта, C — соответственный ущерб, нанесенный «жертвователем» самому себе, а n — число принимающих жертву. Такой принцип позволяет аллелям, способствующим альтруистическому поведению, распространяться в популяции. Но, как отмечает А.В. Марков, «с точки зрения самого „аллеля альтруизма“, в этом никакого альтруизма нет, а есть чистый эгоизм. Этот аллель заставляет своих носителей — то есть организмы — совершать акт альтруизма, но тем самым аллель блюдет свои корыстные интересы. Он жертвует несколькими своими копиями, чтобы дать преимущество другим своим копиям. Естественный отбор — это не что иное, как автоматическое и совершенно равнодушное и бессознательное взвешивание суммы выигрышей и проигрышей для аллеля — для всех его копий вместе — и если выигрыши перевешивают, аллель распространяется». Но родственный отбор — не всегда достаточное средство: ведь для успешного копирования «эгоистичного гена» во многих случаях нужен половой партнер, не состоящий в слишком близком родстве. Тогда могут быть задействованы другие возможности, например, так называемый «реципрокный альтруизм» — особь А делает некоторые действия, выгодные не для нее, а для особи Б, с тем чтобы впоследствии особь Б так же «бескорыстно» помогла особи А. Но и этим возможности «эгоистичного гена» создавать вокруг себя широкий выбор потенциальных половых партнеров не исчерпываются. Как было показано в гл. 4, у многих животных складывается коммуникативная система, которая дает возможность эффективно сосуществовать особям как родственным, так и неродственным, как умеющим, так и не умеющим считать, кто из сородичей кому сколько пользы принес.
В целом, конечно, никто специально не старается трудиться на благо ближнего — особи в своем поведении преследуют свои собственные эгоистические интересы и стремятся устранить конкурентов. Просто у тех видов, которые делают это не слишком эффективно, возникает целый ряд дополнительных преимуществ (прежде всего связанных с выбором оптимального полового партнера). Тем самым оказывается, что создание «супероружия», которое бы позволяло «наиболее приспособленным» особям устранять всех конкурентов раз и навсегда, — с эволюционной точки зрения задача довольно бессмысленная.
У целого ряда животных сформировалась способность жить группами, включающими как родственных, так и не родственных друг другу особей. Если условия окружающей среды неблагоприятны для выживания (вследствие конкуренции с другими группами или высокого уровня активности хищников), отбор на способность к кооперации в группе (в том числе с неродственными особями) усиливается. Так, в одной из популяций шимпанзе в национальном парке Берега Слоновой Кости, соседствующей с крупной популяцией леопардов (самых опасных для шимпанзе хищников), в нескольких группах были зафиксированы случаи усыновления детенышей-сирот. Исследователи специально отмечают, что усыновлять детеныша могут не только родственники или друзья погибшей матери, но и совершенно никак не связанные с ними члены группы (причем не только самки, но и самцы, хотя обычно самцы шимпанзе даже о своих собственных детенышах не слишком заботятся). Очевидно, группы, где достаточно часто появляются склонные к такому альтруизму особи, более эффективно воспроизводят свою численность — действительно, из 18 усыновленных детенышей выжило 10 (более половины), тогда как у сироты, лишенного какой бы то ни было заботы со стороны старших, шансы выжить близки к нулю.
Жить в группе оказывается выгодным не только потому, что это дает возможность кооперироваться при поиске пищи и защите от хищников. Именно от качества группы зависит у таких видов и возможность найти оптимального полового партнера, и выживаемость потомства, и его «воспитание», которое позволит этому потомству в дальнейшем успешно выживать и размножаться. Как пишут М.Л. Бутовская и Л.А. Файнберг, «социальная среда для всех животных является своеобразной „оболочкой“, которая окружает особей и видоизменяет, трансформирует воздействие на них со стороны физической среды». Тем самым «способность к поддержанию взаимоотношений с родственными или неродственными особями являет собой значительное адаптивное преимущество и представляет стратегию, направленную на усиление заботы о потомстве, повышающую вероятность выживания детенышей». И «чем сложнее и гибче социальная организация, тем бóльшую роль она играет в защите особей данного социума». Группы, члены которых были бы ориентированы исключительно на выигрыш у сородичей в конкурентной борьбе и совершенно лишены способности иногда (пусть даже неосознанно) поступиться собственными интересами во благо коллектива, не могли бы иметь эволюционного будущего: из множества нацеленных на победу в конце концов останется только один — победитель. Но, оставшись один, он тем самым лишится адаптивных преимуществ, предоставляемых социальной средой. Поэтому естественный отбор поощряет, с одной стороны, умение особей идти на компромиссы, с другой — разнообразие особей, с тем чтобы каждая из них могла найти свое место в социальной структуре, уменьшая тем самым остроту конкурентной борьбы.
Поскольку ни размеры тела, ни скорость передвижения не могли обеспечить гоминидам безопасность при одиночном образе жизни, особенно в открытых ландшафтах, резонно предположить, что наши предки могли выжить только в достаточно большой группе. Их репродуктивный успех зависел не столько от того, сколько детенышей была способна родить самка, сколько от того, многие ли из них оказывались в состоянии дожить до взрослого возраста, с тем чтобы получить возможность произвести потомство. Когда у ходивших на двух ногах (еще как минимум со времен ардипитеков) гоминид стал увеличиваться мозг, роль социальности сильно возросла, поскольку строение тазовых костей, необходимое для двуногого передвижения, накладывает значительные ограничения на размер головы рождающегося детеныша. М.Л. Бутовская и Л.А. Файнберг полагают, что «уже 1,6 млн. лет назад… длительность беременности стала, по всей видимости, значительно короче общего срока, необходимого для развития мозга, чтобы размеры мозга новорожденного позволяли ему пройти через родовые пути». Рождение слабого, фактически недоношенного детеныша делало необходимым длительную заботу о нем не только матери, но и отца, и других особей. Соответственно, успешно растить детенышей могли лишь те группы, которые были пронизаны прочными социальными связями и в которых был высок уровень внутригрупповой кооперации. В то же время «рождение с маленьким мозгом дает возможность развивать отдельные мозговые структуры после рождения, что позволяет социальной среде принимать значительное участие в формировании мозга» индивида. Чем меньше мозг при рождении (по сравнению с тем, что будет во взрослом состоянии), тем выше роль социальной среды, соответственно, тем больше может быть заложено в мозг после рождения. Чем дольше сроки взросления, тем больше у индивида времени на сравнительно безопасные пробы и ошибки, соответственно, появляется больше шансов для того, чтобы изобрести что-то новое, выучиться чему-то полезному. Развитие мозга и удлинение детства представляют собой процессы с положительной обратной связью: чем больше можно заложить в мозг после рождения, тем более богатую социальную среду может создать соответствующий вид, а чем богаче социальная среда, тем больше она может влиять на развитие мозга. Как пишут Бутовская и Файнберг, «удлинение сроков созревания и связанный с данным процессом рост общих размеров мозга» способствовали возрастанию «значения индивидуальных привязанностей и дружелюбных альянсов». «Увеличение периода созревания и подросткового периода создало дополнительные условия для упрочения связей между родственниками и развития привязанностей между близкими по возрасту индивидами», кроме того, «удлинение подросткового периода в эволюции гоминид вело к повышению роли товарищеских отношений между взрослыми членами группы, способствовало развитию альтруизма и взаимопомощи не только между родственниками, но и между неродственными особями-друзьями. Увеличение же размеров мозга и прямо связанное с ним развитие памяти позволили расширить круг друзей и знакомых». Все эти закономерности наблюдаются и у других приматов, но в гоминидной линии они, судя по всему, получили значительное развитие.
Общественная природа языка подтверждается и тем фактом, что социальная стимуляция играет очень важную роль при обучении языку. Было показано, что у детей-аутистов развитие языка происходит гораздо медленнее и с бóльшими затруднениями, чем у тех, кто реагирует на социальные стимулы нормально.
Но все же социальную функцию языка, видимо, нельзя признать главной движущей силой глоттогенеза. Как показывают наблюдения за ныне живущими приматами, достаточно многие полезные социальные навыки могут существовать и при отсутствии языка. Так, обезьяны умеют поддерживать стабильность группы, мириться (после ссор) и мирить других, а нередко и предотвращать конфликты. Они знают, какие особи с какими состоят в родстве. В эксперименте Верены Дассер яванские макаки отличали фотографии с изображением обезьяны-матери и ее детеныша от снимков, на которых были показаны самка и детеныш, не состоящие в родстве друг с другом. Макаки-резусы, умеющие узнавать сородичей по голосу, четко отличают голоса своих родичей по материнской линии от голосов неродственных особей той же группы. Павианы чакма классифицируют сородичей как по рангу, так и по родству — в экспериментах Д. Чини, Р. Сифарда и их коллег они обращали существенно большее внимание на крики, свидетельствующие о смене иерархических отношений между родственными (по материнской линии) группами, чем в рамках одной такой группы; если павиан стал объектом агрессивного поведения со стороны кого-то из сородичей, он будет избегать не только самого обидчика, но и его родственников.
Обезьяны — не только человекообразные — поддерживают отношения с родственниками и помогают им, могут дружить с неродственными особями, помогать нянчить чужих детенышей в своей группе, заботиться о кормящих матерях. Заботиться о детенышах могут не только самки, но и самцы, причем не только у видов с постоянными парами, но и у видов, для которых типичной формой организации является мультисамцовая группа.
Обезьяны, подобно человеку, «животные политические»: они создают иерархические структуры и добиваются определенного социального статуса; дружественные самцы могут объединиться для свержения доминанта и занятия самого высокого места в иерархии. Даже у видов, живущих мультисамцовыми группами, не принято спариваться с кем попало — для того, чтобы произвести потомство, самцы и самки образуют врéменные брачные пары. У обезьян также имеется отчетливо выраженная, хотя и не абсолютная (впрочем, как и у человека), тенденция избегать инцеста. Опыты Франса де Ваала и Сары Броснан показывают, что обезьянам (причем не только человекообразным) доступны даже такие «высокие материи», как цена или справедливость. В группе капуцинов (см. фото на вклейке) принято нянчить чужих детенышей; для того, чтобы получить такую возможность, надо погруминговать мать малыша. Чем меньше детенышей в группе в данный момент, тем выше цена — дольше необходимый сеанс груминга. Когда же детенышей в группе много, матери соглашаются отдавать их своим подругам за существенно меньшую «плату».
В одном из опытов Ф. де Ваал предлагал капуцинам лакомство за отданный экспериментатору камешек. Но если обезьяна видела, что ей за камешек дают кусочек огурца, а ее соседу — сладкую виноградину, она возмущенно швыряла огурец и отказывалась «работать» в условиях такой вопиющей несправедливости.
Любопытно отметить, что многое из перечисленного и у человека нередко реализуется при помощи не столько языка, сколько невербальной коммуникации. Так, люди оценивают ранг человека по его манере держаться, выбирают полового партнера, ориентируясь в значительной степени на запах и пропорции фигуры, ухаживают при помощи объятий, поцелуев и т.п., утешают, гладя по голове, показывают дружеское расположение при помощи улыбки и т.д., и т.д. Разумеется, люди при этом не думают: «Не стану я ухаживать за этой девушкой — ее запах подсказывает мне, что ее гены плохо сочетаются с моими!» Это решение мозг принимает, не проводя его через сознание, — и человеку просто «почему-то не хочется» ухаживать за соответствующей девушкой, даже если он видит ее многочисленные достоинства. Когда человек чувствует «запах страха» (запах вещества, выделяемого в состоянии испуга), он не осознает этого, но лица на фотографиях «почему-то» существенно (статистически значимо) чаще кажутся ему испуганными — это было установлено посредством эксперимента, в ходе которого людям предъявляли запах страха и фотографии с выражениями лица, промежуточными между удивлением и боязнью. Опыты социопсихологов показывают, что человека, который, входя, занимает место во главе стола, слушаются чаще, чем того, который стремится занять более скромное место (причем, если человек занимает место во главе стола по собственной инициативе, а не по указанию экспериментатора, степень его «убедительности» оказывается выше). Во всех этих (и других подобных) случаях бóльшая часть информации обрабатывается подсознанием, и непосредственный смысл произносимых слов не играет существенной роли.
Кроме всего прочего, естественный отбор — это не только гибель тех, кто хуже приспособлен к выживанию в данных природных условиях, и не только ограничение их участия в размножении. Естественный отбор — это еще и сортировка особей: каждая особь должна найти свое место в иерархии и в ареале. Те особи, которые могут хорошо устроиться в традиционном местообитании, занять место в традиционной группировке, остаются там. Те же, кто не может, оказываются вытесненными на окраинные, менее пригодные для жизни территории. Но если на этих территориях найдутся в достаточном количестве ресурсы (пища, укрытия и т.д.), которые хотя бы некоторые представители вида смогут эффективно использовать, отбор перестанет отбраковывать всех, кто не смог встроиться в традиционную группировку. При достаточном количестве пригодных для использования ресурсов жителей окраин будет становиться все больше, так что с какого-то момента их количество окажется достаточным для формирования группировок и образования брачных пар. Если они сумеют — посредством коммуникации — эти группировки сформировать, существование данного вида на данных территориях станет устойчивым и самовоспроизводящимся. Сформируется новый вариант нормы, сначала опять-таки в рамках видовой нормы реакции. И в традиционных местообитаниях, и в окраинных группировках будут с некоторой частотностью появляться как фенотипы более «традиционные», так и фенотипы, лучше приспособленные для жизни на окраине. Естественный отбор — не в последнюю очередь при помощи коммуникативной системы — будет способствовать аккумуляции первых в традиционном местообитании, а вторых — в окраинном.
Чем больше особей живет «на окраине», тем больше шансов на то, что они смогут создать устойчивые группировки с достаточным разнообразием и тем самым с лучшими возможностями выбора оптимального полового партнера. Генофонд популяции поначалу будет общим, но потом может разделиться — либо из-за каких-то природных причин, которые помешают обмену особями между центром и окраиной, либо вследствие того, что, поскольку многие особи находят себе полового партнера в своей группировке, в разных частях ареала могут накопиться мутации, закрепляющие более высокую вероятность наследования тех признаков, которые необходимы именно в данном местообитании. Эти мутации, которые могут быть незначительны сами по себе, тем не менее могут препятствовать новым поколениям особей встроиться в группировку другого типа местообитания. Кроме того, при приспособлении к новым условиям происходит изменение половых предпочтений: наиболее привлекательными в глазах противоположного пола становятся те, кто лучше всего приспособлен именно к этим условиям. Тем самым половой отбор начинает вместе с естественным работать на разделение ранее единого вида на два.
Одним из недавних примеров такого видообразования может служить разделение рыб-цихлид (род Amphilophus) в никарагуанском озере Апойо на два вида — A. citrinellus и A. zaliosus. Это озеро, представляющее собой залитый водой вулканический кратер и полностью изолированное от других водоемов, образовалось менее 23 тысяч лет назад. Некоторое время спустя, как установили генетики, в озере появилась прародительница всех тамошних цихлид — самка вида A. citrinellus. Ее потомки быстро заселили нишу придонных поедателей водорослей — генетические данные указывают на вспышку численности. Потом численность стабилизировалась — все места, пригодные для нормальной жизни A. citrinellus, оказались уже заняты. С этого момента те особи, которые в силу тех или иных причин оказались несколько мельче других, проиграв конкуренцию более крупным собратьям, стали вытесняться на периферию: они могли плавать в толще воды и питаться насекомыми (которых обычные представители вида A. citrinellus едят редко), но ни к комфортной жизни у дна, ни к спариванию с крупными самками у них доступа не было — в борьбе за существование побеждает сильнейший. Чем больше таких изгоев, тем больше вероятность, что рано или поздно среди них появятся те, чей (случайно сложившийся) фенотип окажется достаточно хорошо приспособленным к окраинной жизни. В данном случае для этого нужно было соединить щуплое телосложение с возможностью охотно и успешно поедать насекомых и со стремлением искать полового партнера не среди представителей «золотого стандарта» A. citrinellus, а среди подобных себе щуплых особей. Те, кому посчастливилось в результате генетических изменений обрести способность передавать по наследству весь этот комплекс признаков одновременно, составили основу нового вида — A. zaliosus. Освободившись от необходимости вести конкурентную борьбу с A. citrinellus, представители нового вида стали быстро заселять озеро — вторая вспышка численности, фиксируемая по популяционно-генетическим данным, продолжается по сей день.
Примерно таким же образом, судя по всему, шел и процесс возникновения человека с его видоспецифической коммуникативной системой — языком.
С наступлением эпохи более холодного и сухого климата перед приматами открывается новая возможность — огромные (и все увеличивающиеся) пространства, занимаемые полуоткрытыми (редколесье) и открытыми (саванна) ландшафтами. До миоцена (а именно в конце миоцена появляются первые представители клады человека), судя по палеонтологическим данным, ничего похожего на степи или саванны в Африке не было — саванны возникают в процессе совместной эволюции «злаков и крупных травоядных млекопитающих». Их начинают осваивать самые разные группы животных — таким образом в интервале 3–2,5 млн. лет назад появляются десятки новых видов грызунов и крупных копытных. Для привычных к условиям влажного тропического леса приматов такие местообитания представляются скорее неудобными, поскольку там хуже, чем в лесу, работают привычные стратегии добывания пищи, поиска укрытий и т.п. Но по количеству кормовых ресурсов эти места не настолько безнадежны, чтобы особи, не сумевшие встроиться в лесные группировки, немедленно погибали. Пищи в редколесье и даже в саванне вполне достаточно — надо только суметь ею воспользоваться. Но условия ее поиска в такого рода мозаичных ландшафтах совершенно не таковы, как во влажных тропических ландшафтах, где обитали предки гоминид. Источники пищи существенно меняются в разные сезоны — во влажное время года в изобилии имеется растительная пища, термиты, птичьи яйца и т.д., в сухой сезон можно есть семена и корни трав, ловить рыбу в высыхающих водоемах, питаться мясом — либо охотясь на травоядных, сосредотачивающихся в это время года около источников воды, либо поедая трупы павших животных. Длина влажного и сухого сезонов в разные годы неодинакова, что делает жизнь в саванне еще более разнообразной и непредсказуемой. Кроме того, в саваннах, где было много травянистой растительности и, соответственно, много травоядных животных, «увеличилось видовое разнообразие и общая численность хищников. Такое положение вещей с неизбежностью приводило к ситуации, основным плюсом которой было появление новых источников пищи, а основным недостатком — повышение риска встречи с хищником», и это тоже было очень существенным фактором, к которому гоминиды должны были адаптироваться.
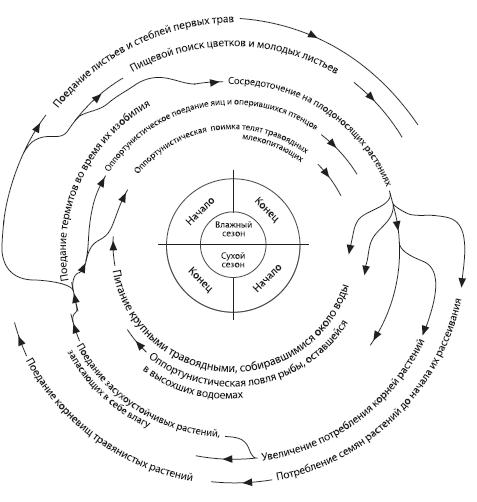
Рис. 5.4. Модель оптимизации и приспособления к условиям для пищевого поиска гоминид на протяжении сезонного цикла (с учетом сезонных изменений доступности пищевых ресурсов), предложенная Р. Фоули.
Вообще, открытые ландшафты, по-видимому, дают больше, чем лесные, разнообразных условий среды, к которым можно приспосабливаться. Если, например, рассмотреть эволюцию в семействе полевок, то можно увидеть, что род рыжих полевок, живущих в лесу, содержит сравнительно немного видов, и все они довольно архаичны, то есть сформировались достаточно давно и с тех пор не менялись. Напротив, серые полевки, освоившие луговые местообитания, демонстрируют целый спектр сравнительно молодых видов — это значит, что им приходилось то и дело приспосабливаться к меняющимся условиям среды.
О том, что экологическая ниша, которую начали осваивать гоминиды, была для них новой, свидетельствует, в частности, полиморфизм ранних представителей клады человека (независимо от того, являются ли найденные останки действительно принадлежащими столь многим разным видам или даже родам или в ряде случаев демонстрируют лишь разнообразие локальных вариаций в рамках одного рода или вида). Возникновение любого нового таксона «до некоторой степени аналогично проникновению на мало заселенные острова» (т.е. в новую экологическую нишу), и на начальных этапах его существования имеет место «бурная дивергентная эволюция», причем ранние члены таксона даже на низких таксономических уровнях резко различаются между собой, «порой по признакам, которые позже будут характеризовать таксоны гораздо более высокого ранга» (так называемый «закон архаического многообразия»). Движение от разнообразия к единообразию в процессе происхождения человека иллюстрируют в том числе и следующие факты. У Homo habilis наблюдается значительная вариативность по объему мозга (от 500 до 800 см3) — и на следующую ступень эволюции «переходят» лишь обладатели самого большого мозга (в итоге у архантропов он достигает более 900 см3). Точно так же у Homo heidelbergensis сильно варьировали возможности слухового анализатора (см. рис. в гл. 3) — и для перехода на следующую ступень эволюции были отобраны обладатели лишь одного из возможных вариантов. Видимо, отбор шел на приспособленность к определенным поведенческим свойствам, а сами свойства появлялись в ответ на некий эволюционный вызов. Примерно так же происходило образование нового вида тлей в опытах Георгия Христофоровича Шапошникова: при насильственной пересадке в другую экологическую нишу (на другое растение) наблюдалось сначала повышение вариативности, а потом (за 170 поколений) из этого множества вариантов отобрались наиболее приспособленные, и появился новый вид (не только отличающийся по своему строению, но и не скрещивающийся с видом-предком).
О том, какую именно нишу стали осваивать далекие предки человека, существуют разные мнения. Одни исследователи полагают, что гоминиды занимались поеданием падали — в это время сильно уменьшилось число видов крупных гиен, так что невостребованных трупов должно было быть в достатке. Кроме того, гоминиды за счет того, что отсутствие волос, обилие потовых желез и выпрямленная походка предохраняли их от перегрева, могли быть активны в жаркие дневные часы, избегая тем самым конкуренции с другими любителями мяса (и хищниками, и падальщиками), которые в это время отдыхают. По мнению других, они могли охотиться. Но в любом случае, «наблюдаемые увеличения общих размеров тела, объема мозга, продолжительности жизни не могли иметь места без смены пищевой стратегии, а смена пищевой стратегии означает переход в новую экологическую нишу».
Для такого перехода необходимо развитие нужных специализаций. Но природа, как уже говорилось, не производит неограниченного спектра форм, из которых естественный отбор мог бы выбирать наилучшие — эволюция обычно идет по пути развития того, что у организмов уже существует. Для нее оказывается существенно проще все более и более усиливать уже существующую тенденцию к той или иной специализации, чем отказываться от нее и переходить к специализации совершенно иного типа, поэтому, например, среди кошачьих появляются все более и более эффективные хищники, но не возникает травоядных форм. Эволюция приматов не является исключением из общего правила.
Как отмечают Бутовская и Файнберг, предковые для гоминид формы скорее всего были всеядны и не имели выраженных специализаций к какому-либо определенному типу добычи пищи. У них «не было ни быстроты стайных хищников, ни их зубов, ни когтей для охоты», ни таких развитых анатомических приспособлений для питания грубой растительной пищей, как у копытных. Конкурировать с другими представителями фауны наши предки могли прежде всего за счет способности быстро адаптироваться к окружающей среде посредством поведения. Поведенческое приспособление имеет то преимущество перед приспособлением анатомо-физиологическим, что осуществляется в более короткие сроки, позволяет более оперативно реагировать на изменения окружающей среды, быстро осваивать новые условия (и тем самым расширять свою экологическую нишу).
Рис. 5.5. Чтобы научиться лакомиться сливками, никакие мутации не нужны. Как показали Дэвид Шерри и Беннет Галеф, не нужно даже наблюдать, как проклевывают крышку: одного вида бодро скачущего поблизости сородича достаточно, чтобы побудить синицу продырявить крышку и добраться до сливок.
В нынешнее время мы можем наблюдать это на примере освоения городов различными видами птиц. Например, грач (Corvus frugilegus) освоил города Приднестровья за 15 лет, вяхирь (Columba palumbus) за 10 лет смог сформировать жизнеспособную популяцию во Львове. Поведение городских птиц сильно отличается от поведения их диких сородичей: грачи зимой кормятся на тротуарах, почти под ногами у прохожих, подобно голубям и воробьям, ястребы-тетеревятники (Accipiter gentilis) осваивают охоту на чердаках, выслеживают добычу в густых сумерках и даже «мышкуют» среди травы. Подобные нетипичные для диких птиц варианты поведения распространяются за время гораздо меньшее, чем понадобилось бы в случае, если бы эти поведенческие характеристики возникали в результате мутаций. Как отмечает орнитолог Вадим Вадимович Корбут, «формирование урбанизированных популяций серой вороны и кряквы в Москве в 60–80-е годы привело к нарастанию их численности на 2–3 порядка при жизни особей-основателей». Впрочем, следует отметить, что эти элементы поведения именно нетипичны, а не вовсе невозможны для соответствующих видов — было показано, что они входят в видовую норму реакции. Подобного же рода быстрое распространение поведенческих признаков наблюдали в сороковые годы в Англии на примере синиц-лазоревок (Parus caeruleus), за считанные годы научившихся открывать закупоренные фольгой молочные бутылки. Широко известны наблюдения за японскими макаками острова Кошима, самостоятельно додумавшимися до того, что клубни батата (сладкого картофеля) можно мыть в ручье. Сначала этот способ очистки пищи от грязи применяла только одна самка Имо — изобретательница метода, потом постепенно он распространился по всей группировке.
Рис. 5.6. А потом Имо догадалась, что клубень станет вкуснее, если надкусить его и окунуть в соленую морскую воду. Другие макаки, конечно же, собезьянничали.
Отметим специально, что рассмотренные изменения в поведении — это не результат мутаций, возникших у единичной особи и затем распространившихся: для мутационного сценария необходимо существенно больше эволюционного времени.
Такого рода поведенческое приспособление может иметь важные эволюционные последствия: оно запускает механизм, известный как «эффект Болдуина» (по имени американского психолога Джеймса Марка Болдуина, сформулировавшего в 1896 году эту гипотезу). Выглядит он следующим образом. Пусть у каких-то особей данного вида есть в арсенале некоторая модель поведения, которая приносит им выгоду — помогает более эффективно находить пищу, избегать опасностей и т.д. Это значит, что их генетические характеристики, как минимум, не препятствуют реализации этой модели. Обладатели этих генетических характеристик оставят больше потомства, значит, в следующем поколении особей, которые обладают способностью к этому поведению, будет больше, и снова максимум потомства оставят именно они. В наибольшем эволюционном выигрыше оказываются те, чьи гены не только не препятствуют полезному поведению, но прямо ему способствуют, причем вовсе не обязательно превращать это поведение в инстинкт — достаточно просто уметь (быть генетически предрасположенным) быстро и надежно ему обучаться. Подчеркнем специально, что, когда в регуляции развития особи достаточно большое участие принимает внешняя среда, генетически передается не само сложное поведение (в частности, коммуникативное), а предрасположенность к нему, к тем или иным реакциям индивида на явления окружающей действительности — т.е. в конечном счете к тому, чтобы некоторые связи между нейронами устанавливались быстрее и были более прочными, чем другие.
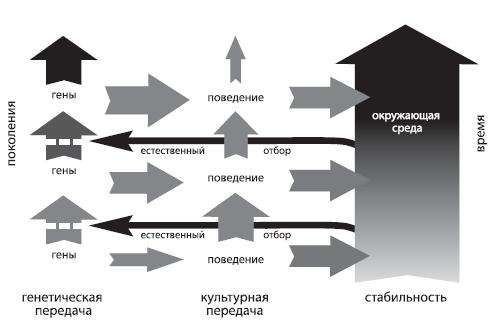
Рис. 5.7. Эффект Болдуина
Когда преимущества, которые дает то или иное поведение, достаточно серьезны, отбор благоприятствует тем популяциям, где особи, способные к нему, появляются чаще. Именно так, скорее всего, происходило и формирование языка — через постепенное накопление (сначала — в рамках видовой нормы реакции) тех, кто лучше умел доносить свои мысли до окружающих, и тех, кто был наиболее успешен в понимании. Если бы языковая способность возникла в результате мутации у единичной особи, язык не смог бы сформироваться — так же, как не складывался язык в никарагуанской школе для глухих, пока число стремящихся общаться между собой воспитанников не перевалило за две сотни.
Если то или иное поведение окажется полезным, будут закрепляться мутации, способствующие совместному наследованию всего того комплекса признаков, который обеспечивает данное поведение. Будут поддерживаться отбором и те мутации, которые будут изменять анатомию организма в сторону, благоприятствующую данному поведению. Например, успехи в ухаживательном поведении, включающем демонстрацию развернутого веером хвоста, формируют в конечном итоге хвост, весьма выигрышно выглядящий в развернутом виде. Вероятно, такого же рода эволюцию можно предполагать и для языка: если, например, стремление произносить звуковые сигналы максимально внятно и разборчиво приносило ощутимые выгоды в деле выживания и размножения, то отбором закреплялись такие анатомические свойства, которые максимально облегчали особям эту задачу (пусть даже в ущерб каким-то другим жизненным интересам), — и в итоге люди получили свою опущенную гортань. Если выгодно было уметь передавать максимум информации за один раз, закрепились мутации, позволявшие (пусть с большей тратой ресурсов) более эффективно управлять дыханием, и т.д.
Этот механизм эволюции отличается от широко распространенных представлений, согласно которым сначала возникает мутация, а потом организмы пытаются извлечь из нее какую-то пользу. В данном случае, наоборот, сначала формируются те или иные поведенческие навыки, и лишь потом те особенности организма, которые позволяют реализовывать их наилучшим образом, закрепляются генетически (что обеспечивает эффективное наследование поведения без прямого закрепления в генах конкретных поведенческих моделей). Согласно этому сценарию, мутации — необходимый компонент эволюционного процесса, но для каждого отдельного эволюционного события генетическое закрепление тех или иных характеристик фенотипа — не начальная, а, скорее, завершающая стадия. Какие именно фенотипические характеристики могут возникать в ходе эволюции подобным образом — вопрос сложный, и окончательного ответа на него в науке пока нет, но то, что в эволюции поведения эффект Болдуина играет значительную роль, по-видимому, факт. А следовательно, для происхождения языка такой эволюционный механизм представляется более вероятным, нежели обратный (сначала — мутации, потом поведение).
Способности к поведенческому приспособлению имеются у многих животных, но человекообразные обезьяны продвинулись в этом значительно дальше других. Они способны не только осознать один элемент ситуации как средство обработки другого (например, ручей как средство для мытья клубней или камень как средство для того, чтобы расколоть орех), они — по крайней мере, шимпанзе — могут целенаправленно изготавливать орудия труда или переносить требующую обработки пищу в удобное для обработки место (см. гл. 3). Вообще, для видов, не имеющих выраженной специализации к определенному типу пищи, «характерны более разнообразный поведенческий репертуар и способность к полифункциональному применению элемента поведения, обобщению типичных для вида элементов в необычные комплексы-комбинации». Кроме того, всеядность имеет то преимущество перед растительноядностью, что позволяет иметь менее массивные кости челюстей и черепа в целом, а это, в свою очередь, позволяет увеличить объем мозга (а тем самым и возможность строить и хранить в памяти большее количество поведенческих программ) без создания дополнительных нагрузок на опорно-двигательный аппарат, поскольку «при уменьшении объема кости на единицу объем мозга увеличивается на две единицы» (при сохранении той же массы).
Основная специализация приматов в природе — понимание причинно-следственных связей, умение строить поведенческие программы, исходя не из каких-то предзаданных шаблонов, а из представления о внутреннем устройстве наличной ситуации. Именно поэтому в условиях эксперимента они оказываются в состоянии догадаться, что высоко висящий банан можно достать, построив пирамиду из ящиков или соединив две палки, огонь спиртовки, мешающий добраться до лакомства, можно залить водой и т.д.
У людей способность понимать причинно-следственные связи развита неизмеримо сильнее, чем у обезьян, следовательно, ее развитие занимало важное место в эволюции человека. В мозгу эту способность обеспечивают префронтальные формации коры. Так, обезьяна с поврежденной префронтальной корой не способна понять, что, если пищу на ее глазах переложили из ящика А в ящик В, то искать ее в ящике А бессмысленно, обезьяна же с неповрежденной корой легко справляется с подобными задачами. Человек (но не обезьяна) справляется и с задачей выбрать меньшее лакомство, если известно (из предыдущего опыта), что выбранное будет отдано другому (см. гл. 2). Иными словами, префронтальные формации коры обеспечивают возможность делать выводы из нескольких посылок, реагировать на стимул не непосредственно, а с учетом разных привходящих обстоятельств. Умения такого рода чрезвычайно важны для всеядного существа (например, они позволяют, увидев дерево с незрелыми плодами, вернуться к нему через некоторое время, когда плоды созреют). Они же играют значительную роль в обеспечении функционирования языка, позволяя понимать сообщения, содержащие несколько утверждений. Кроме того, префронтальные отделы коры помогают планировать сложное поведение (в том числе коммуникативное), диктуют, что замечать, а что игнорировать, — это позволяет, в частности, опознавать одинаковые слова, произнесенные разными людьми, с разной громкостью, скоростью и т.п., как одно и то же слово. В гоминидной линии идет постоянное увеличение размеров (как абсолютных, так и относительных) префронтальной коры.
Согласно гипотезе американского антрополога Дуайта Рида, именно увеличение префронтальной коры явилось одним из важнейших факторов глоттогенеза. Дело в том, что в одном из ее участков (поля 9 и 46 по Бродману) расположен называемый «центральный исполнительный компонент» (англ. central executive component) рабочей памяти, позволяющий удерживать внимание на нескольких объектах одновременно. Человек может оперировать одновременно примерно семью понятиями (точнее, 7±2), а шимпанзе — всего лишь двумя-тремя. Как показывают наблюдения приматологов, шимпанзе сравнительно легко обучаются колоть орехи, манипулируя одновременно двумя предметами (орехом и камнем-молотком), но с гораздо бóльшим трудом овладевают умением колоть орехи, требующим координированных действий с тремя предметами — орехом, камнем-молотком и камнем-наковальней. Увеличение объема рабочей памяти (англ. ST-WMC, short-term working memory capacity) позволяет человеку, в отличие от обезьян, строить и понимать предложения, состоящие из многих слов, а также овладевать правилами их построения.
Главным эволюционным «козырем» гоминид был, пользуясь терминологией А.А. Зубова, «комплекс гоминизации». В него входят «высокоразвитый мозг, анатомические особенности руки и возможность ее освобождения для использования, а затем изготовления орудий труда и охоты, стереоскопическое зрение, сложное групповое поведение, включая высокий уровень организации стада, достаточно развитые средства коммуникации, взаимодействие между индивидуумами (кооперацию), склонность к манипулированию предметами, гибкость поведения, высокий удельный вес поведенческих адаптаций в общей системе приспособительных реакций, повышенная роль прижизненного опыта в организации поведения и, соответственно, — возрастающая роль индивидуума в группе». Как пишут Бутовская и Файнберг, «дифференциация поведения особей на индивидуальном уровне, связанная с уникальностью их прижизненного опыта в сочетании с индивидуальными психическими задатками… способствовала повышению пластичности функционирования группы в целом и обеспечивала определенный запас адаптаций к изменяющимся условиям среды. Тенденция к индивидуализации поведения прослеживается в сравнительном ряду приматов».
Появление новой экологической ниши привело к тому, что при сортировке по поведенческим стратегиям «естественный отбор на „линии шимпанзе“ стал усиливать качества, повышающие приспособленность к жизни в лесу небольшими коллективами, в то время как на линии гоминизации (очеловечивания) требовалось развитие таких качеств, как организация, сплоченность сравнительно большой группы, способной жить как в лесу, так и на границе саванны, более совершенные средства общения — коммуникации между членами группы, координация их действий». «Комплекс гоминизации» (как целое) становится «объектом действия движущего естественного отбора, определяющего направление дальнейшего развития и видообразования». У тех же, кто был лучше адаптирован для жизни в лесу, наоборот, развиваются специализации к лесной жизни — стопы, пригодные для лазанья по деревьям, руки, удобные не только для лазанья, но и для хождения с опорой на костяшки пальцев, зубная система, приспособленная к специфически лесной диете.
Первоначально между прежними и новыми биотопами шел постоянный обмен генетическим материалом (ср.: «сочетание интеграции на генетическом и дифференциации на экстрагенетическом уровнях позволяет популяциям одного вида приматов занимать различные экологические ниши и создает возможность для широкого расселения вида, освоения новых территорий, позволяет одновременно с этим сохранить видовое единство»). Но климат становился все более холодным и сухим, саванны вытесняли леса, и постепенно территории, на которых жили гоминиды, отделялись друг от друга. Обмен особями (и, соответственно, генами) прекращался, и в разделившихся популяциях накапливались разные генетические изменения. У обитателей более лесных биотопов они были направлены на все лучшее приспособление к жизни в лесу (затем — в редколесье и т.д.) и соответствующему питанию, у обитателей более новых биотопов — на дальнейшее совершенствование поведения, развитие способности постигать причинно-следственные связи и реагировать на события окружающей действительности «с опережением». Соответственно, чем более гоминиды продвигались в саванны, тем больший спрос возникал на развитие коммуникативной системы.
Жизнь на границе более закрытого и более открытого ландшафта благоприятствует тому, чтобы особи, не забывая старых способов добычи пищи, поиска укрытий и т.п., осваивали новые. Количество потенциально возможных для вида поведенческих программ таким образом растет. Еще более оно возрастает с появлением орудий — помимо навыков, позволяющих добыть пищу при помощи орудий, полезно помнить навыки, позволяющие добыть ее «голыми руками». Кроме того, полезно представлять себе, как эти орудия делаются, где добывается для них материал и т.д. Еще один фактор роста числа поведенческих моделей — переход к более активному употреблению мяса. Мясо стало важным источником энергии, и, следовательно, репертуар пищедобывательных стратегий расширился — независимо от того, какими конкретно способами это мясо добывалось (была ли это охота, поедание падали или же комбинация того и другого). При этом «не исключено, что изменение рациона и укрупнение мозга усиливали друг друга и способствовали развитию более сложного социального поведения, что вызывало дальнейшее совершенствование тактики пищевого поиска и диеты. А это, в свою очередь, благоприятствовало дальнейшей эволюции головного мозга». С одной стороны, увеличивающийся мозг дает возможность поддерживать более развитую коммуникативную систему, с другой — появляется способность принимать во внимание все большее и большее количество факторов внешней среды, все тоньше различать детали, позволяющие отличить одну ситуацию от другой, и на развитие коммуникативной системы возникает спрос. Можно, конечно, этого и не делать, но тогда для выживания необходимо встроиться в более «традиционную» группировку — и продолжать существование до тех пор, пока старые местообитания еще остаются.
Процесс возникновения человеческого языка недоступен непосредственному наблюдению, поэтому судить о нем можно лишь по косвенным данным. В качестве одного из источников таких данных нередко называют развитие языка у ребенка. Стадии, выделяемые в овладении языком, представляются легко сопоставимыми либо с последовательно сменявшими друг друга видами гоминид, либо с палеолитическими индустриями. Основанием для этого служит так называемый «основной биогенетический закон» Эрнста Геккеля — онтогенез (т.е. развитие единичной особи) есть сжатое и сокращенное повторение хода филогенеза (т.е. развития таксона).
Но все же, по-видимому, не следует прямо проецировать становление языка у ребенка на происхождение человеческого языка в целом. Биологам хорошо известно, что закон Геккеля представляет собой лишь частный, хотя и довольно широко распространенный случай. Он выполняется только для отдельных органов при их преобразовании по способу анаболии (т.е. добавления еще одной стадии развития после всех тех, что имелись у вида-предка). Как было показано А.Н. Северцовым, эволюционируют не только фенотипы взрослых особей, но и их онтогенезы. Как пишет Н.Н. Иорданский, «изменчивость организмов проявляется на всех стадиях индивидуального развития и служит материалом для действия естественного отбора». Онтогенез человека достаточно сильно изменился по сравнению с онтогенезом обезьян. Если детеныш обезьяны от рождения до взрослого состояния проходит две стадии — младенчество и детство, то у человека таких стадий осмысленно выделять четыре. Первая из них (от рождения до двух с половиной — трех лет) несколько перестроена по сравнению с обезьяньим младенчеством, вторая (до момента смены зубов) возникает в человеческой линии около 2 млн. лет назад (у обезьян этой стадии нет), третья (до начала пубертатного периода) соответствует детству (= подростковому возрасту) обезьян, но также претерпела значительные перестройки, и наконец, четвертая стадия (собственно подростковая, примерно до 19 лет), судя по палеонтологическим данным, появляется у неоантропов, а у Homo erectus и более ранних гоминид отсутствует. Эти периоды различаются не только по тому, какие телесные структуры формируются в течение каждого из них, но и по тому, какие изменения претерпевает когнитивная сфера, а также какими аспектами языка овладевает ребенок (см. гл. 1).
У человека, который усваивает язык, изначально (до появления речи) присутствует целый ряд необходимых для этого когнитивных установок, отсутствующих у обезьян, — стремление интерпретировать звуки, которые произносят другие люди, как знаки, желание узнавать названия различных объектов, установка на кооперативность в поведении вообще и в коммуникации в частности и т.д. (см. гл. 2). Соответственно, на каких-то этапах эволюции человека они должны были войти в программу его онтогенеза. О перестройке программы индивидуального развития свидетельствует и то, что период овладения звуком (лепет) в ней предшествует периоду, когда ребенок начинает понимать, что окружающие имеют цели и намерения.
Еще одно весьма существенное отличие онтогенеза и филогенеза языка состоит в том, что человек выучивает язык в общении с теми, кто его уже знает (и это является необходимым условием, см. гл. 1; даже если родители — носители пиджина, все равно есть хотя бы один настоящий язык, которым они владеют в совершенстве, — их родной язык). Как показывают наблюдения психолингвистов, диалоги детей со взрослыми более чем вдвое (в среднем) длиннее, чем диалоги детей с детьми (лучше всего детям удается поддерживать беседу, когда они вступают в разговор взрослого с ребенком на пару лет старше их, — в этом случае их реплики не сбивают разговор с обсуждаемой темы, а самих реплик может быть больше, чем обычно бывает в разговоре с этим же взрослым или с этим же ребенком).
Надо сказать, что взрослые, общаясь с ребенком, обычно не только не хвалят его за грамматически правильные предложения, но даже не всегда исправляют его ошибки, — нередко они обращают внимание не на грамотность, а на истинность сказанного, ср. такие диалоги:
(1) РЕБЕНОК: Mamma isn’t boy, he a girl («Мама не мальчик, он девочка»; в первой части пропущен артикль, во второй — глагол-связка.)МАМА: That’s right. («Правильно».)
(2) РЕБЕНОК: And Walt Disney comes on Tuesday («А Уолта Диснея показывают по вторникам»; грамматических нарушений нет.)МАМА: No, he does not. («Нет, не так».)
Тем не менее языковое поведение взрослых при общении с ребенком имеет целый ряд специфических черт, призванных способствовать овладению языком. «По сравнению с разговорами между взрослыми, речь родителей, обращенная к детям, медленнее, в ней более утрирована высота тона, она больше говорит о происходящем здесь и сейчас, и она более грамматически правильна». Четкие паузы на синтаксических границах, отсутствие семантически сложных слов и конструкций также облегчают для ребенка понимание обращенных к нему высказываний. Как пишет С.Н. Цейтлин, при общении с детьми «мы стараемся строить предложения правильно, планируем их структуру, не перестраиваемся на ходу, не обрываем посередине и т.п.», — в отличие от того, что часто можно наблюдать при обычных «взрослых» разговорах. «Еще одно существенное свойство речи, адресованной ребенку, — частые повторы, обычно сопровождаемые перестановкой компонентов предложения: „Мячик упал у Мишеньки. Дай мне мячик. Красивый мячик“. Это помогает ребенку членить высказывание, выделяя в нем слова». Кроме того, «матери повторяют многое из сказанного ребенком, корректируя эти фрагменты в звуковом и грамматическом отношении», ср. примеры, приводимые С.Н. Цейтлин:
(1) МАМА: Кто это?РЕБЕНОК: ‘ХУГУМАМА (с утвердительной интонацией): Хрюша.
(2) РЕБЕНОК (про котенка): Она не хочет гладить.МАМА: Не хочет, чтобы ты гладил его?
Подобного рода корректирующие реплики можно наблюдать и в разговорах взрослых, ср., например: «Мы ужинали с ним со среды на пятницу». — «Вы хотели сказать: со среды на четверг». Но, разумеется, в диалогах с детьми они встречаются гораздо чаще.
Кроме того, взрослый, разговаривая с ребенком, следит за проявлениями его внимания и при необходимости может скорректировать свое речевое поведение. Если же ребенок слышит только не обращенную к нему речь взрослых (такое случается, например, со слышащими детьми глухих родителей, которые включают своему малышу телевизор), это не способствует усвоению им языка.
В некоторых культурах взрослые мало общаются с маленькими детьми, но зато с ними могут говорить другие дети, и тогда именно они будут обеспечивать малышам этот особый регистр языка, называемый иногда «материнским языком» (англ. motherese). Дети достаточно рано начинают понимать специфику этого регистра — как отмечает психолингвист Ревекка Марковна Фрумкина, «уже четырехлетние дети в разговорах с двухлетними пользуются более короткими словами, чем в разговорах со сверстниками или со взрослыми».
Все это говорит о том, что онтогенез речевой коммуникации никак нельзя считать прямым повторением филогенеза. Какие-то параллели между выучиванием языка детьми и возникновением языка в процессе эволюции человека проводить, безусловно, можно — но эти параллели, скорее, сами нуждаются в доказательствах, чем могут служить таковыми.
Еще один вопрос, который обычно задают, когда речь идет об эволюции человеческого языка, — это вопрос о том, почему не появляется язык у человекообразных обезьян, хотя они достаточно умны, пользуются орудиями (и обучают этому детенышей), умеют их изготавливать, демонстрируют феноменальные способности к овладению языками-посредниками в языковых проектах. Правда, согласно старинному поверью, говорить обезьяны все-таки умеют, но никогда не делают этого в присутствии людей, чтобы те не заставили их работать. Если же серьезно, ответ скорее всего таков: потому, что имеющейся у них системы коммуникации вполне достаточно для успешного выживания и размножения. Как уже говорилось (см. гл. 3), исследователи обнаружили у шимпанзе в природе 39 различных моделей поведения, которые являются обычными в одних группах, но отсутствуют в других, — в это число входит и использование орудий, и груминг, и ухаживание. И это всё, чего смогли достичь шимпанзе за 6–7 миллионов лет своего существования как отдельного вида. У самых «примитивных» людей (что бы ни вкладывать в понятие «примитивности») культурно-специфичных практик (даже без учета языка) насчитывается гораздо больше. Видимо, у шимпанзе жизнь все же более однообразна, чем у людей, и поэтому нет потребности в увеличении количества сигналов. А вот в условиях неволи количество сигналов может возрастать: добиваясь от людей ласки, лакомства и т.п., шимпанзе весьма активно изобретают новые элементы поведения, и, «поскольку такие инновации почти всегда подкрепляются, частота проявления у „авторов“ растет вместе с числом подражателей — наиболее предприимчивых сородичей». В природных же условиях подкрепления изобретаемых сигналов в большинстве случаев нет, и они угасают. Один такой случай приводит Дж. Гудолл: в Гомбе самка-подросток Фифи вдруг стала встряхивать кистями рук — этот жест она употребила, угрожая самке старшего возраста. В этот момент с Фифи была еще одна, более молодая самка — Гилка. На следующей неделе Гилка в сходной ситуации употребила этот жест. Какое-то время он продержался в репертуаре обеих, а потом сошел на нет.
Некоторые из обезьян, как видно по данным экспериментов, проявляют больше способностей к овладению языком, некоторые — меньше, но жесткого отбора, при котором первые получали бы заметные преимущества перед вторыми, в природе нет. Точно так же и среди людей встречаются, например, великолепные имитаторы — они могут изобразить кудахтанье курицы, скрип двери и даже пение канарейки. Но отбора, который бы давал таким людям преимущество перед людьми, лишенными этого таланта, нет. Такие фенотипы, не выходящие за рамки нормы реакции, появляются с определенной частотой, но, поскольку необходимость в этом признаке отсутствует, закрепления его в генах не происходит.
Рассуждая о происхождении человеческого языка, трудно обойти вопрос о том, было ли оно неизбежностью или случайностью. На мой взгляд, оно было неизбежным — в том смысле, что являлось закономерным логическим продолжением той адаптации к мыслительной деятельности, на путь которой вступили приматы. У гоминид развивались не анатомо-физиологические приспособления к определенным условиям окружающей среды, а орудийная деятельность, способность делать выводы (подтверждаемая увеличением префронтальных отделов коры) и — вследствие группового образа жизни — коммуникативная система. Таким образом, отбор благоприятствовал развитию (в числе прочего) успешности коммуникации, что и привело в итоге к появлению вида, высокоспециализированного в этой области, — человека разумного. Но и роль случайности в процессе глоттогенеза не следует недооценивать — ведь если бы на Земле сложились другие экологические условия, столь далеко зашедшая специализация, возможно, и не потребовалась бы.

