Книга: Рассказ предка. Паломничество к истокам жизни
Назад: Рандеву № 6 Обезьяны Нового Света
Дальше: Рандеву № 7 Долгопяты
Рассказ Ревуна
Новые гены появляются не из воздуха. Они образуются при дупликации прежних генов. После этого каждый из них идет своим путем, приобретая мутации и подвергаясь действию естественного отбора и дрейфа генов. Обычно мы не видим, как это происходит, но, подобно следователям на месте преступления, можем попытаться воссоздать произошедшее на основе улик. Замечательный пример – гены, ответственные за цветовое зрение.
Млекопитающие очень долго были ночными существами. День принадлежал динозаврам, у которых, если судить по ныне живущим родственникам, было отличное цветовое зрение. Также не без оснований можно предположить, что цветовое зрение имелось и у далеких предков млекопитающих – звероподобных рептилий (тероморфов), населявших Землю до динозавров. Однако за время, которое млекопитающие провели во тьме, их глаза должны были научиться улавливать каждый доступный фотон. Неудивительно, что способность различать цвета они почти утратили (о причинах мы поговорим в “Рассказе Слепой пещерной рыбы”). До сих пор у большинства млекопитающих (даже тех, которые вернулись к дневному образу жизни) слабое цветовое зрение – двухроматическое. Речь о количестве типов светочувствительных клеток – колбочек – в сетчатке. У человека и остальных узконосых обезьян, а также у обезьян Старого Света, три типа колбочек: красные, зеленые и синие. То есть у нас зрение трихроматическое. Однако есть данные, что мы заново приобрели третий тип колбочек после того, как его утратили наши ночные предки. У большинства других позвоночных, например рыб или рептилий (но не млекопитающих!), зрение трихроматическое (имеются колбочки трех типов) или тетрахроматическое (четыре типа). У птиц и черепах зрение бывает еще сложнее. Что касается обезьян Нового Света, особенно ревунов, то они в особенном положении.
Есть указания на то, что австралийские сумчатые, в отличие от большинства млекопитающих, обладают хорошим трихроматическим зрением. Кэтрин Аррес и ее коллеги, открывшие его у поссумов-медоедов и австралийских сумчатых землероек (такое зрение было найдено и у кенгуру-валлаби), предположили, что австралийские (но не американские) сумчатые сохранили древний светочувствительный пигмент рептилий, утраченный остальными млекопитающими. Однако в целом у млекопитающих все-таки самое слабое среди позвоночных цветовое зрение. Большинство млекопитающих если и различает цвета, то примерно на уровне страдающих дальтонизмом людей. Любопытные исключения из этого правила можно найти среди приматов. Неслучайно они чаще других млекопитающих используют для брачной демонстрации яркие цвета.
Австралийские сумчатые, в отличие от нас, видимо, никогда не теряли цветовое зрение. Мы же, судя по своим родственникам среди млекопитающих, скорее всего, не унаследовали от рептилий трихроматическое зрение, а заново изобрели его. Причем два раза: первый раз это сделали обезьяны Старого Света и человекообразные обезьяны, а второй – ревуны (только они, а не все обезьяны Нового Света). Цветовое зрение ревунов похоже на цветовое зрение человекообразных обезьян, однако отличается от него достаточно, чтобы понять: возникло оно независимо.
Что же такого хорошего в цветовом зрении, что трихроматизм возник независимо у обезьян Старого и Нового Света? Основная гипотеза гласит, что цветовое зрение связано с питанием фруктами. На фоне однотонной зеленой листвы фрукты выделяются. Это, в свою очередь, не случайно. Фрукты скорее всего приобрели яркие цвета для привлечения поедающих плоды животных, например обезьян, которые играют важную роль в распространении семян. Кроме того, трихроматическое зрение помогает находить на фоне темной листвы более молодые и мясистые листья (которые бывают бледно-зелеными, иногда даже красными). Впрочем, последнее вряд ли идет на пользу растениям.
Цвет завораживает нас. Слова, обозначающие цвета – первые прилагательные, которые узнают дети, и именно эти слова они раньше всего пытаются связать с каким-нибудь подходящим существительным. Но мы редко вспоминаем, что цвета и оттенки, которые мы видим – это ярлыки, которые наше сознание навешивает на участки спектра, слегка различающиеся длиной электромагнитных волн. Красный свет соответствует длине волны около 700 нм, фиолетовый – около 420 нм. При этом видимое излучение в этих рамках составляет ничтожную часть спектра. Ведь длина волны может варьировать от нескольких километров (некоторые радиоволны) до долей нанометра (гамма-излучение).
Все глаза на планете устроены так, чтобы воспринимать волны тех длин, в которых наша звезда ярче всего и которые проходят сквозь земную атмосферу. Однако способность глаза к восприятию электромагнитных волн ограничена биохимическими системами, необходимыми для улавливания размытого диапазона излучения. Законы физики устанавливают для видимого диапазона длин волн более четкие границы. Ни одно животное не воспринимает всю инфракрасную область спектра. Ближе всего подошли к этому гремучие змеи. У них на голове есть ямки, которые хотя и не могут создавать сфокусированное изображение в инфракрасном диапазоне, дают змеям некоторую направленную чувствительность к теплу, исходящему от добычи. Также ни одно животное не может воспринимать весь диапазон ультрафиолетовой части спектра, хотя некоторые животные, например пчелы, видят в ультрафиолете немного лучше нас. Зато пчелы не видят “нашего” красного цвета: для них он инфракрасный. В целом для животных “свет” – это узкий спектр электромагнитных волн между короткими волнами ультрафиолетового излучения и длинными – инфракрасного. Различие между пчелами, людьми и змеями лишь в том, где проходят границы воспринимаемого спектра.
Более узки эти границы для каждого типа светочувствительных клеток сетчатки. Одни колбочки немного лучше воспринимают красный участок спектра, другие – синий. Цветовое зрение представляет собой сопоставление данных, полученных от различных колбочек, и качество такого зрения сильно зависит от того, сколько типов колбочек в сетчатке. В сетчатке животных с дихроматическим зрением – два типа колбочек, трихроматическим зрением – три, тетрахроматическим зрением – четыре. Каждая колбочка характеризуется кривой чувствительности, у которой есть пик и не очень симметричные “угасающие” концы. За пределами, очерченными этой кривой, клетка “слепа”.
Допустим, колбочка имеет пик в зеленой части спектра. Значит ли это, что такая клетка посылает сигналы в мозг лишь тогда, когда она “смотрит” на зеленый объект, например на траву или бильярдный стол? Безусловно, нет. Просто клетке необходимо больше красного света (в этом случае), чтобы выдавать импульсы той же интенсивности, как при заданном количестве зеленого света. Такая клетка будет одинаково реагировать как на яркий красный свет, так и на тусклый зеленый. Нервная система может определить цвет объекта лишь путем сопоставления одновременно поступающих импульсов по меньшей мере от двух клеток, чувствительных к разным участкам спектра. Каждая клетка при этом “контролирует” другую. Если же клеток не две, а три, представление о цвете окажется еще полнее.
Цветное телевидение и компьютерные мониторы работают в трехцветной системе, потому что были разработаны для трихроматических глаз. В обычном мониторе каждый пиксель состоит из трех точек, расположенных настолько близко друг к другу, что глаз не может их различить. Каждая точка всегда светится одним цветом. Посмотрев на экран под достаточно большим увеличением, мы увидим одни и те же три цвета – обычно красный, зеленый и синий, хотя иногда могут использоваться и другие комбинации. Любой оттенок можно получить, регулируя интенсивность свечения трех основных цветов. Должно быть, черепах с их тетрахроматическим зрением наши телевизоры разочаровывают.
Сопоставляя интенсивность импульсов всего от трех типов колбочек, наш мозг может воспринимать огромный диапазон оттенков. Но большинство плацентарных млекопитающих обладает дихроматическим зрением: в их сетчатке колбочки двух типов. У колбочек одного типа пик восприимчивости в фиолетовой области спектра (в некоторых случаях – в ультрафиолетовой), у второго типа – где-то между зеленой и красной областями. У нас, животных с трихроматическим зрением, пик “коротковолновых” колбочек находится между фиолетовой и синей областями спектра. Такие колбочки называют синими… Другие два типа наших колбочек – так называемые зеленые и красные. Правда, даже у красных колбочек пик приходится скорее на желтоватую, чем на красную часть спектра. Но в целом кривая их чувствительности сдвинута в красную часть спектра. Так что, несмотря на то, что пик в желтой области, эти колбочки все равно генерируют сильный импульс в ответ на красный свет. Поэтому, если вычесть интенсивность импульса от зеленых колбочек из интенсивности импульса красных колбочек, мы получим особенно сильный сигнал в ответ на красный свет.
Кроме колбочек, в сетчатке еще есть палочки – светочувствительные клетки, которые отличаются от колбочек формой и особенно эффективны ночью. В цветовом зрении эти клетки не участвуют, и мы больше не будем о них говорить.
Химия и генетика цветового зрения довольно хорошо изучены. Главными молекулярными акторами здесь выступают опсины. Это молекулы белка, которые в колбочках и палочках выполняют функцию зрительных пигментов. Каждая молекула опсина связана с одной молекулой ретиналя – химического соединения, которое является производным витамина А . Молекула ретиналя скручена в петлю, которая встраивается в молекулу опсина. При попадании фотона с подходящей длиной волны узел распрямляется. Это служит сигналом для клетки: она посылает нервный импульс, который говорит мозгу: “Вижу свет своего типа”. Тогда молекула опсина связывается с новой скрученной молекулой ретиналя, поступающей из внутриклеточных запасов.
Не все молекулы опсина одинаковы. Опсины, как и все белки, кодируются генами. Различия в ДНК приводят к образованию опсинов, чувствительных к разным цветам, и это служит генетической основой дихроматического и трихроматического зрения. А поскольку каждая клетка организма имеет полный набор генов, различия между красными и синими колбочками не в том, какие гены у них есть, а в том, какие гены работают. На этот счет есть правило: каждая колбочка включает лишь один тип генов.
Гены, кодирующие наши зеленый и красный опсины, очень похожи друг на друга и находятся на Х-хромосоме (половая хромосома, которая у женщин есть в двух копиях, у мужчин – в одной). Ген, отвечающий за образование синего опсина, немного от них отличается и находится не на половой хромосоме, а на одной из обычных хромосом, которые называют аутосомами (в нашем случае это седьмая хромосома). Наши зеленые и красные клетки явно образовались в результате недавней дупликации гена, который, в свою очередь, задолго до этого образовался в результате дупликации гена синего опсина. Тип зрения животного – дихроматический или трихроматический – зависит от того, сколько разных генов опсинов у него в геноме. Если в геноме животного есть, например, гены синего и зеленого опсинов, но отсутствует ген красного опсина, такое животное будет дихроматом.
Прежде чем заняться ревунами и историей приобретения ими трихроматического зрения, попытаемся понять, как устроено странное для нас дихроматическое зрение остальных обезьян Нового Света. Кстати, дихроматическое зрение есть у некоторых лемуров, но не у всех обезьян Нового Света (у ночных обезьян зрение монохроматическое). Так что пока забудем о ревунах и других необычных видах.
Во-первых, исключим из обсуждения “синий” ген, который находится на аутосомах всех особей независимо от пола. С “красными” и “зелеными” генами на Х-хромосоме все сложнее и гораздо интереснее. На каждой Х-хромосоме есть только один локус, в котором может находиться красный или зеленый аллель. Поскольку у самки две Х-хромосомы, она может обладать и красным, и зеленым аллелями одновременно. Однако у самца с его единственной Х-хромосомой может быть лишь один аллель: красный либо зеленый. Таким образом, типичный самец обезьяны Нового Света должен обладать дихроматическим зрением, потому что у него колбочки лишь двух типов: синие плюс красные либо зеленые. По нашим меркам, все эти самцы – дальтоники. Однако это дальтоники двух типов: у одних самцов в популяции нет зеленого опсина, у других – красного. Синий есть у всех.
Самкам теоретически повезло больше. Поскольку у них две Х-хромосомы, они могут оказаться счастливыми обладателями красного аллеля на одной хромосоме и зеленого аллеля – на другой (плюс, конечно, синий аллель). Такая самка будет обладать трихроматическим зрением. У менее удачливой самки может оказаться два красных или два зеленых аллеля, и тогда ее зрение будет дихроматическим. По нашим меркам такие самки тоже будут дальтониками и тоже, как и в случае с самцами, дальтониками двух типов.
Поэтому популяции обезьян Нового Света, например тамаринов или беличьих обезьян (саймири), представляют собой странную смесь. Все самцы и некоторые самки обладают дихроматическим зрением, то есть они дальтоники, причем двух типов. Некоторые самки – но не самцы – обладают полноценным трихроматическим зрением, которое предположительно похоже на наше. Эксперименты, в ходе которых тамарины искали еду в замаскированных коробках, показали, что особи с трихроматическим зрением делают это успешнее, чем особи с дихроматическим зрением. Возможно, что, выходя на поиски пищи, стаи обезьян Нового Света полагаются на счастливых обладательниц трихроматического зрения. С другой стороны, возможно, обладатели дихроматического зрения, поодиночке или в компании с “дальтоником” другого типа, могут обладать неожиданными преимуществами. Говорят, во время Второй мировой войны в экипажи бомбардировщиков включали одного дальтоника, потому что он мог различить некоторые виды маскировки лучше товарищей-трихроматов. Эксперименты подтверждают, что люди с дихроматическим зрением распознают некоторые типы камуфляжа, способные обмануть людей с полноценным зрением. Так что, возможно, группа обезьян, состоящая из обладателей трихроматического зрения и “дальтоников” двух типов, найдет больше фруктов, чем группа, состоящая лишь из животных с трихроматическим зрением. Звучит надуманно, но смысл в этом есть.
Гены красного и зеленого опсинов у обезьян Нового Света представляют собой пример полиморфизма. Это одновременное существование в популяции двух или более альтернативных версий гена. При этом ни одна из версий не должна быть слишком редкой, то есть представлять собой результат недавней мутации. Известный принцип эволюционной генетики гласит: полиморфизм, подобный этому, не возникает без причины. Если бы он возникал просто так, в конце концов обезьян с красным аллелем стало бы больше, чем обезьян с зеленым аллелем, – или наоборот. Какой именно аллель получил бы преимущество, мы не знаем, но вероятность того, что оба оказались бы одинаково полезны, очень мала. Так что в итоге худший из них обязательно был бы отсеян.
Существование в популяции стабильного полиморфизма говорит о том, что он нужен. Но зачем? На этот счет существует две гипотезы, и каждая в нашем случае вполне применима. Это частотно-зависимый отбор и гетерозиготное преимущество. Явление частотнозависимого отбора наблюдается тогда, когда преимущество получает более редкий тип – просто потому, что он более редкий. При этом по мере вымирания менее предпочтительного типа он становится все более редким – и тогда получает преимущество. Допустим, что обезьяны с красным аллелем особенно хорошо различают красные фрукты, а с зеленым аллелем – зеленые. Если в популяции преобладают обезьяны с красным аллелем, все красные фрукты скоро будут съедены, и тогда преимущество получит одинокая обезьяна с зеленым аллелем, которая умеет находить зеленые фрукты. И наоборот. Хотя эта история звучит не слишком правдоподобно, она служит примером условий, необходимых для сохранения в популяции обоих типов. Эти необходимые условия в целом напоминают историю с летчиками-дальтониками.
Теперь обратимся к явлению гетерозиготного преимущества. Классическим примером выступает серповидноклеточная анемия у человека. Ген серповидноклеточной анемии плох тем, что у индивидов, имеющих две его копии (то есть у гомозигот по этому гену), эритроциты деформируются и становятся похожими на серпы. Такие люди страдают от тяжелой формы анемии. Однако у этого гена есть и плюсы: люди, имеющие лишь одну его копию (то есть гетерозиготы по этому гены), обладают устойчивостью к малярии. В тех районах, где распространена малярия, польза от этого гена перевешивает его вред, и ген серповидноклеточной анемии сохраняется в популяции, несмотря на пагубное воздействие на тех, кому не посчастливилось иметь две его копии. Профессор Джон Моллон и его коллеги многое сделали для понимания полиморфной системы цветового зрения у обезьян Нового Света. Они полагают, что гетерозиготного преимущества, которым обладают самки с трихроматическим зрением, достаточно для сохранения красных и зеленых аллелей в популяции. Но еще лучшим примером в этом смысле является ревун.
Ревуны ловко пользуются преимуществами обоих механизмов полиморфизма, объединив их на одной хромосоме благодаря удачной транслокации (это особый вид мутации). При транслокации происходит перенос участка хромосомы на другую хромосому – или в другое место на той же хромосоме. Похоже, именно это произошло у предка ревунов. В результате оба аллеля – и красный, и зеленый – оказались рядом на одной Х-хромосоме. Даже если этот предок был самцом, указанная транслокация послужила отправной точкой для эволюционного развития трихроматического зрения. Со временем мутантная Х-хромосома распространялась в популяции, и теперь она есть в геноме у всех ревунов.
Ревунам было легко проделать этот эволюционный фокус, потому что генофонд популяции обезьян Нового Света уже содержал все три аллеля гена опсина. Просто в геноме каждой обезьяны, за исключением некоторых удачливых самок, было только два аллеля. Между тем у человекообразных обезьян и обезьян Старого Света трихроматическое зрение возникло по-другому. Наши дихроматические предки не делились на два типа, то есть в их популяции не было полиморфизма. Есть основания полагать, что удвоение гена опсина на Х-хромосоме наших предков было настоящей дупликацией. Первый мутантный предок оказался обладателем двух тандемных (то есть расположенных рядом) копий одного и того же аллеля – например зеленого. Поэтому он, в отличие от мутантного предка ревунов, не стал внезапным обладателем трихроматического зрения. У него было обычное дихроматическое зрение, обусловленное одним синим и двумя зелеными аллелями. Трихроматическое зрение у обезьян Старого Света формировалось постепенно, в ходе последующей эволюции. Поколение за поколением естественный отбор благоприятствовал расхождению цветовой чувствительности двух опсиновых аллелей на Х-хромосоме, что привело к образованию зеленого и красного аллелей.
При транслокации переносу подвергается не один ген, а сразу несколько. И иногда его спутники – соседи по хромосоме, перемещающиеся вместе с ним на новую хромосому, – могут рассказать кое-что интересное. Это именно такой случай. Ген Alu известен как “мобильный генетический элемент”: это короткий вирусоподобный участок ДНК, который размножается в геноме как своего рода паразит, использующий клеточный механизм репликации ДНК. Участвовал ли Alu в перемещении опсина? Похоже, что так. Изучив геном, мы можем найти неопровержимые доказательства. На обоих концах дуплицированного участка находятся гены Alu. Возможно, дупликация была случайным побочным эффектом репродукции “паразитического” элемента. В геноме некоей эоценовой обезьяны рядом с геном опсина имелся паразитический элемент. Во время самовоспроизведения он случайно реплицировал гораздо больший, чем нужно, участок ДНК и так создал предпосылку к появлению у нас цветового зрения. Тут возникает искушение сделать вывод, что если геномный паразит случайно оказал нам услугу, значит, геномы предоставляют убежище паразитам в надежде на гипотетическую пользу. Но это не так. Естественный отбор так не работает.
Ошибки копирования, подобные описанной, могут иногда происходить без участия Alu. Например, когда две Х-хромосомы объединяются перед кроссинговером, они с некоторой вероятностью могут сделать это неправильно. И тогда вместо правильного выравнивания (красный аллель на одной хромосоме встает напротив такого же красного аллеля на другой хромосоме) красный аллель оказывается, например, напротив зеленого. Происходит это из-за сходства аллелей. Если за такой ошибкой следует кроссинговер, он получается “неравным”: на одной хромосоме может оказаться лишний зеленый ген, а на другой – ни одного. Но даже если кроссинговер не происходит, может наблюдаться “конверсия генов”. При конверсии генов короткая последовательность на одной хромосоме превращается в соответствующую последовательность на другой. При ошибочном выравнивании хромосом участок красного аллеля может заменить соответствующий участок зеленого аллеля – и наоборот. Таким образом, к “красно-зеленому” дальтонизму может привести как неравный кроссинговер, так и конверсия генов при ошибочном выравнивании хромосом.
Красно-зеленым дальтонизмом (то есть неспособностью различать красный и зеленый цвета) чаще страдают мужчины, чем женщины. Ну, не то чтобы они страдали, однако это причиняет определенное неудобство: дальтоникам предположительно недоступны некоторые эстетические удовольствия. У мужчин, в отличие от женщин, нет запасной Х-хромосомы, которая могла бы заменить дефектную. Никто не знает, видят ли дальтоники кровь и траву такими же, как мы. Возможно, это зависит от человека. Мы знаем лишь, что людям с красно-зеленым дальтонизмом предметы цвета травы кажутся примерно такими же, как предметы цвета крови. В человеческой популяции дихроматический дальтонизм наблюдается примерно у 2 % мужчин. Кстати, не путайте дихроматический дальтонизм с другими видами красно-зеленого дальтонизма, которые встречаются гораздо чаще (примерно у 8 % мужчин). Таких людей называют аномальными трихроматами: хотя генетически они являются трихроматами, опсины одного из трех типов у них не работают.
Неравный кроссинговер не всегда плох. Бывает, на некоторых Х-хромосомах оказывается более двух генов опсина. Дополнительный аллель почти всегда зеленый, а не красный. Рекорд – двенадцать дополнительных зеленых аллелей, расположенных тандемом на Х-хромосоме. Мы не знаем, лучше ли у таких индивидов зрение. Однако известно, что не все “зеленые” аллелли идентичны друг другу, – поэтому теоретически возможно не только трихроматическое, но и тетрахроматическое, даже пентахроматическое зрение. Не знаю, пробовал ли кто-нибудь это проверить.
Из моих слов можно заключить, что приобретение нового опсина в результате мутации автоматически приводит к улучшению цветового зрения. Однако ясно, что различия в чувствительности колбочек бесполезны до тех пор, пока мозг не умеет понимать, от какого из типов колбочек приходит сигнал. Если бы это достигалось с помощью жесткой генетической схемы, в которой каждая колбочка связана с конкретной клеткой головного мозга, система вполне могла бы работать, но не могла бы справляться с мутациями, возникающими в сетчатке. Да и как? Откуда мозг может “знать”, что в сетчатке вдруг появился новый опсин, чувствительный к другому цвету, и что в какой-то группе колбочек “включился” ген этого нового опсина?
Думаю, самый убедительный ответ таков: мозг учится. По-видимому, он сравнивает силу импульсов от различных групп колбочек в сетчатке и может “заметить”, что одна группа клеток генерирует более сильный импульс при виде помидоров и клубники, вторая – при виде неба, третья – травы. Разумеется, это догадка. Но я думаю, что примерно такой механизм лежит в основе способности нервной системы быстро приспосабливаться к генетическим изменениям в сетчатке. Мой коллега Колин Блейкмор считает, что такого рода проблемы возникают всякий раз, когда нервной системе нужно приспособиться к изменениям на периферии.
Генная дупликация очень важна. Гены красного и зеленого опсинов явно произошли от предкового гена, который скопировал себя в новое место на Х-хромосоме. А перед этим в результате сходной дупликации синий ген на аутосоме отделился от предка красного и зеленого аллелей на Х-хромосоме. Часто гены, находящиеся на разных хромосомах, принадлежат к одному и тому же “семейству генов”. Такие семейства возникают в результате древних дупликаций с последующим расхождением функций. В нескольких исследованиях было показано, что типичный ген человека подвергается дупликации со средней вероятностью 0,1–1 % за 1 млн лет. Дупликация ДНК может быть поэтапной или мгновенной: скажем, когда в геноме распространяется новый активный ДНК-паразит, например Alu, или когда дупликации подвергается геном целиком. (Удвоение всего генома распространено у растений; кроме того, считается, что при зарождении позвоночных это произошло по меньшей мере дважды.) Как бы то ни было, случайные дупликации ДНК – один из основных источников новых генов. В ходе эволюции меняются не только гены в геномах – меняются и сами геномы.
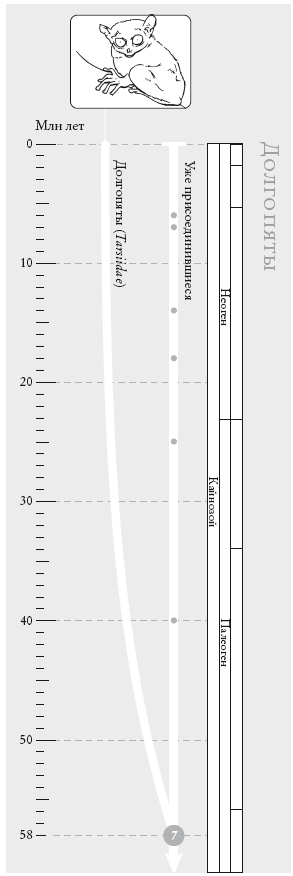
Долгопяты. Согласно последним морфологическим и молекулярным данным, пять видов долгопятов образуют сестринскую группу по отношению к обезьянам Старого и Нового Света и к человекообразным обезьянам. Прежде долгопятов объединяли с лемурами.
На рис.: филиппинский долгопят, или сирихта (Tarsius surichta).
Назад: Рандеву № 6 Обезьяны Нового Света
Дальше: Рандеву № 7 Долгопяты

