Книга: Рассказ предка. Паломничество к истокам жизни
Назад: Рассказ Тасманийца[2]
Дальше: Архаичный Homo sapiens
Рассказ Митохондриальной Евы
Между родословными генов и людей есть заметная разница. В отличие от человека, у которого всегда два родителя, у гена “родитель” лишь один. Каждый из ваших генов достался вам либо от матери, либо от отца; только от одного из четырех бабушек и дедушек; только от одного из восьми прабабушек и прадедушек, и так далее. Но когда люди составляют родословную, они считают своими предками в равной степени обоих родителей (четырех прародителей, восьмерых прапрародителей и так далее). “Генеалогия людей”, таким образом, гораздо запутаннее “генеалогии генов”. В определенном смысле ген следует одним-единственным путем из множества пересекающихся дорожек, лежащих в основе человеческой родословной. Подобно генам ведут себя наши фамилии, но не люди. Наша фамилия следует лишь одной ветви раскидистого генеалогического древа и передается по мужской линии. ДНК же, за исключением двух случаев, не женоненавистница: родословная генов с равной вероятностью прослеживается и по мужской, и по женской линии.
Генеалогические древа европейских королевских семей входят в число человеческих родословных, задокументированных наилучшим образом. В Саксен-Кобург-Готской династии обратите особое внимание на принцев Алексея, Вальдемара, Генриха и Руперта (см. рис.). “Родословную” одного из их генов проследить особенно легко, потому что (к несчастью для них) ген был дефектным. Из-за него четыре принца и многие другие члены злополучной семьи приобрели гемофилию. Это заболевание крови передается по Х-хромосоме. У мужчин лишь одна Х-хромосома, которую они наследуют от матери. У женщин Х-хромосом две – по одной от каждого родителя. Женщины болеют гемофилией, лишь если унаследовали дефектную версию гена и от матери, и от отца (гемофилия – “рецессивное” заболевание). Мужчины же болеют, если дефектный ген несет единственная “незащищенная” Х-хромосома. Таким образом, от гемофилии страдает ничтожно мало женщин, при этом многие женщины являются ее носителями и с пятидесятипроцентной вероятностью передают ее ребенку. Такие женщины надеются, что родят дочь, но и в этом случае остается риск того, что гемофилия перейдет к внукам. Если больной гемофилией мужчина проживет достаточно долго для того, чтобы иметь детей, он не сможет передать дефектный ген сыну (мужчины никогда не получают Х-хромосому от отца), но обязательно передаст его дочери (женщины всегда получают единственную Х-хромосому отца). Помня эти правила и зная, у каких мужчин из королевской династии была гемофилия, можно проследить путь дефектного гена.

Наследование гемофилии в Саксен-Кобург-Готской династии.
Судя по всему, первой мутантным геном обзавелась королева Виктория. Это не был ее муж Альберт, потому что его сын, принц Леопольд, был гемофиликом, а сыновья получают А-хромосому не от отца. Ни один из родственников Виктории в побочной линии не страдал от гемофилии. Ошибка при копировании могла произойти в яйцеклетке ее матери, Виктории Саксен-Кобург-Заальфельдской, или (это вероятнее по причинам, о которых рассказывает Стив Джонс в книге “Язык генов”), “в августейших яичках ее отца, Эдуарда Августа, герцога Кентского”.
Хотя сами родители Виктории не были носителями дефектного гена и не страдали от гемофилии, очевидно, у одного из них имелся ген (строго говоря – аллель), который стал предмутантным “родителем” гена гемофилии. Мы можем попробовать проследить судьбу этого гена королевы Виктории до его мутации. Для нас несущественно, что у Виктории копия гена была повреждена, а у ее предшественников – нет. Изучая “родословную” гена, мы игнорируем все его проявления, кроме тех, которые делают его “видимым”. Ген должен был появиться задолго до Виктории, но видимый его след утерян, потому что в то время он еще не был геном гемофилии. Мораль в том, что у каждого гена один “родительский” ген, даже если в результате мутации они стали непохожими. Точно так же у родительского гена есть лишь один родитель, а у того – свой, и так далее. Эти рассуждения могут показаться странными, но помните: мы ищем предков. Именно так выглядит поиск предка с точки зрения гена.
В “Рассказе Тасманийца” мы ввели понятие “последний общий предок” (ПОП), альтернативное понятию “сопредок”. Я хочу зарезервировать “сопредка” за последним общим предком всей родословной (людей или организмов). Поэтому, говоря о генах, я буду говорить о ПОП. Два или более аллелей у разных особей (или даже у одной) обязательно имеют ПОП. Это предковый ген, и каждый из этих аллелей – его копия (возможно, мутантная). ПОП генов гемофилии принцев Вальдемара и Генриха Прусских находился на одной из двух А-хромосом их матери Ирены Гессен-Дармштадтской. Когда она была еще в утробе, две копии гена гемофилии, носителем которого она была, разделились и попали последовательно в две ее яйцеклетки, из которых получились ее несчастные сыновья. Эти гены, в свою очередь, имеют ПОП с геном гемофилии русского царевича Алексея (1904–1918): это ген, носителем которого была их бабушка – Алиса Гессенская. И, наконец, ПОП генов гемофилии этих четырех принцев – тот самый ген, с которого мы начали разговор: мутантный ген королевы Виктории.
У генетиков есть термин для ретроспективного анализа генов: коалесцентный. Если посмотреть в прошлое, линии этих двух генов в некотором роде сходятся (коалесцируют) в одной точке. Если же из этой точки снова взглянуть вперед, мы увидим, как две копии родительского гена передаются двум потомкам. Точка схождения этих линий и есть ПОП. В любой генной “родословной” много точек схождения. Линии генов гемофилии Вальдемара и Генриха сходятся в точке гена ПОП – их матери Ирены. Эта линия затем соединяется с ветвью, ведущей к царевичу Алексею. И, как мы видели, окончательно линии генов гемофилии сходятся на Виктории.
Она, ко всему, их последний общий генеалогический предок (“прародитель”), то есть их сопредок. Но это совпадение. Если бы мы выбрали другой ген (например, ген цвета глаз), его путь по генеалогическому древу был бы другим, и гены сходились бы в более далеком предке. Если бы мы выбрали ген карих глаз у принца Руперта и ген голубых глаз у принца Генриха, точка их схождения была бы по меньшей мере столь же отдаленной, как разделение предкового гена цвета глаз на две формы. А это событие таится во мраке веков. У каждого фрагмента ДНК есть “родословная”, которую можно узнать примерно так, как родословную человека – по метрическим книгам.
Мы можем сделать это даже для двух идентичных генов одного человека. У принца Чарльза голубые глаза. Это рецессивный признак, и, значит, у него было два аллеля голубых глаз. Эти два аллеля должны сходиться, но где и когда, мы не знаем. Это могло быть несколько веков и даже тысячелетий назад, но в случае принца Чарльза два аллеля голубых глаз, возможно, сходятся в королеве Виктории. Она предок Чарльза сразу по двум линиям: через короля Эдуарда VII и принцессу Алису Гессен-Дармштадтскую. Согласно этой гипотезе, ген голубых глаз Виктории копировался дважды. Эти копии достались, соответственно, нынешней королеве (правнучке Эдуарда VII) и ее мужу, принцу Филиппу (правнуку Алисы). Таким образом, две копии одного гена Виктории, вполне возможно, встретились на двух разных хромосомах принца Чарльза. Вообще-то с некоторыми его генами это произошло наверняка, пусть даже не с геном голубых глаз. И независимо от того, сходятся ли два гена голубых глаз в королеве Виктории или более далеком предке, у этих двух генов в любом случае есть ПОП. Неважно, говорим мы о двух генах одного человека (Чарльз) или о двух генах двух людей (Руперт и Генрих): логика не меняется. В отношении любых двух аллелей (одного или разных людей) закономерен вопрос: когда и в ком сошлись эти гены в прошлом? Этот вопрос можно задать относительно любых трех (или другого числа) генов в популяции, если они занимают в геноме одно и то же положение (локус).
Если заглянуть еще глубже в прошлое, этот вопрос можно задать и касательно пар генов в различных локусах, потому что гены в процессе дупликации порождают гены в других локусах. К этому мы вернемся в “Рассказе Ревуна” и “Рассказе Миноги”.
Степень родства у людей можно представить как своего рода решение, принятое большинством генов. Некоторые ваши гены “голосуют”, например, за королеву в качестве близкой родственницы. Другие полагают, что вы ближе к более далеким родственникам (иногда даже к представителям других видов). Если “опросить” разные участки ДНК, то выяснится, что у всех собственное представление о прошлом. Полную картину можно получить, опросив большое количество генов. Но нужно быть осторожным с генами, расположенными близко на хромосоме. Чтобы понять, почему, нужно узнать кое-что о рекомбинации при образовании сперматозоида или яйцеклетки.
Хромосомы при рекомбинации обмениваются случайными участками тождественной ДНК. В среднем на одну хромосому человека приходится всего один-два таких обмена (при образовании сперматозоида – меньше, при образовании яйцеклетки – больше, причем неизвестно, почему). С поколениями все больше участков хромосомы меняются местами. Чем ближе расположены два участка ДНК на хромосоме, тем меньше шансов, что между ними произойдет обмен, и выше вероятность, что они будут наследоваться вместе.
Поэтому при “подсчете голосов” генов следует помнить, что чем ближе друг к другу гены на хромосоме, тем вероятнее, что у них общая история. Такие гены при голосовании поддерживают друг друга. В крайнем варианте участки ДНК оказываются настолько сильно связаны, что весь фрагмент – гаплотип – ведет себя как целое. Единогласие в “парламенте генов” нарушают лишь два его члена – и не потому, что у них особое видение истории, а потому, что их чаще других использовали для улаживания биологических “дебатов”. Оба придерживаются сексистских взглядов: один передается лишь через женские тела, а второй никогда не бывал вне мужского. Это два главных исключения из принципа равномерного генного наследования.
Как и фамилия, К-хромосома (та ее часть, которая не участвует в рекомбинации) передается лишь по мужской линии. К-хромосома содержит генетический материал, который фактически переключает эмбрион на мужской тип развития. Митохондриальная ДНК, напротив, передается исключительно по женской линии (хотя и не отвечает за женский путь развития эмбриона: у мужчин тоже есть митохондрии, но они не передают их потомкам). На Великом историческом рандеву мы увидим, что митохондрии – это крошечные тельца в клетках, потомки некогда свободноживущих бактерий, которые около 2 млрд лет назад на особых правах поселились внутри клеток. Там они размножались бесполым способом – простым делением. Многие свои бактериальные свойства они утратили, как и большую часть ДНК, но того, что сохранилось, оказалось достаточно для генетиков. Митохондрии составляют независимую линию генетической преемственности, не связанную с главной ядерной линией, о которой мы привыкли думать как о “наших” генах.
Скорость мутирования К-хромосомы делает ее удобным инструментом для исследований современных популяций. В ходе одного исследования ученые собрали образцы ДНК К-хромосомы из точек на прямой, проходящей через современную Великобританию. Результаты показали, что К-хромосомы англосаксов двигались из Европы на запад и довольно резко остановились у границы Уэльса. Нетрудно представить, почему ДНК, которая переносится мужчинами, не является показательной для остальной части генома. Вот более очевидный пример: корабли викингов, которые несли груз К-хромосом (и других генов), распространили его на огромной территории. Современное распределение генов К-хромосомы викингов указывает, что путешествовать им нравилось сильнее, чем остальным генам викингов, предпочитавшим странствиям родной очаг.
Митохондриальная ДНК тоже может показать неплохие результаты, особенно в отношении древних событий. Если сравнить нашу митохондриальную ДНК, можно будет сказать, как давно жил общий митохондриальный предок. А поскольку мы получаем митохондрии от матерей, а те – от бабушек по материнской линии, прабабушек по материнской линии и так далее, мы можем выяснить, когда жил наш последний предок по материнской линии – наша праматерь. То же можно сделать и для У-хромосом. Это поможет узнать, когда жил наш последний предок по мужской линии – наш праотец. Впрочем, это не так-то просто сделать по техническим причинам. Прелесть У-хромосомной и митохондриальной ДНК в том, что они не “загрязнены” половым смешением.
Митохондриальную праматерь человечества, которая соответствует общему предку всех людей по материнской линии, иногда называют “Митохондриальной Евой”. А общего предка человечества по мужской линии можно назвать “У-хромосомным Адамом”. У всех мужчин есть У-хромосома Адама (креационисты, молчите). Если бы передача фамилий всегда соответствовала нынешним правилам, все мы носили бы фамилию Адама. Впрочем, в этом случае фамилия, наверное, утратила бы смысл.
Ева – великая искусительница, так что будем начеку. Во-первых, важно понимать, что Адам и Ева – лишь два представителя множества последних общих предков. Конечно, это предки особенные: мы встретим их, если двинемся по генеалогической схеме соответственно от матери к матери или от отца к отцу. Но есть масса других способов передвижения по генеалогическим схемам: от отца матери к отцу его матери, от матери матери к отцу ее отца, и так далее. В конце каждой линии стоит свой ПОП.
Во-вторых, Ева и Адам не были парой. Если они и были знакомы, то случайно. Возможно, их разделяли десятки тысяч лет. Есть основания полагать, что Ева жила раньше Адама. Репродуктивный результат у мужчин варьирует больше, чем у женщин: у некоторых женщин может быть впятеро больше детей, чем у других, а у самых успешных мужчин – в сотни раз больше, чем у неудачников. Мужчина с большим гаремом легко может стать общим предком человечества. Женщине же, поскольку ей не так просто завести настолько обширную семью, для совершения такого подвига необходимо большее число поколений. И действительно, самые точные пока “молекулярные часы” оценивают возраст Евы примерно в 140 тыс. лет, Адама – всего около 60 тыс. лет.
В-третьих, Адам и Ева – это переходящее почетное звание, а не имена конкретных людей. Если завтра умрет последний представитель некоего богом забытого племени, эстафетная палочка Адама или Евы может оказаться на несколько тысяч лет впереди. Это касается и остальных ПОП у основания деревьев других генов. Чтобы понять, почему это так, предположим, что у Евы было две дочери, одна из которых стала прародительницей тасманийских аборигенов, а вторая – остальной части человечества. Также допустим, и не без оснований, что ПОП по женской линии, объединяющий “остальную часть человечества”, жил 10 тыс. лет спустя после того, как вымерли остальные побочные линии, идущие от Евы – кроме тасманийцев. И получается, что когда умерла Труганини, последняя тасманийка, звание Евы немедленно помолодело на 10 тыс. лет.
В-четвертых, в своей эпохе Адам и Ева особенно не выделялись. В отличие от легендарных тезок, Митохондриальная Ева и Е-хромосомный Адам не были одиноки. У них были друзья и знакомые, и каждый из них, возможно, имел немало половых партнеров, от которых тоже могли остаться потомки. Единственное, что выделяет их – у Адама в итоге оказалось огромное количество потомков по мужской линии, а у Евы – по женской. Возможно, вместе они оставили столько же потомков, сколько все их современники.
Когда я это писал, мне прислали документальный фильм Би-би-си “Родина” (Motherland) – “невероятно трогательный” и с “красивейшим, незабываемым сюжетом”. Героями фильма были трое “черных” (почему здесь кавычки, я объясню в “Рассказе Кобылки”), семьи которых переехали с Ямайки в Великобританию. Их ДНК сравнивали с образцами ДНК из международных баз данных, чтобы установить, из какой части Африки вывезли их предков. Продюсерская компания организовала слезное воссоединение героев с давно утраченной африканской родней. В исследовании использовались Е-хромосомная и митохондриальная ДНК: их легче изучать, чем остальные гены. К сожалению, продюсеры не признались, какие ограничения имеет избранный метод. Они намеренно ввели в заблуждение и героев, и их “родственников” (несомненно, у телевизионщиков были веские причины), благодаря чему сцена воссоединения получилась гораздо эмоциональнее, чем полагалось.
Когда Марк, которому дали родовое имя Каигама, приехал в племя канури в Нигере, он считал, что “возвращается” в земли “своего народа”. Болу приветствовали на острове у побережья Гвинеи восемь женщин из племени буби, с которыми у нее совпала митохондриальная ДНК. Бола говорила:
Это как кровь соприкасается с кровью… Это как семья… Я плакала, мои глаза были полны слез, мое сердце билось. Я думала только об одном – я возвращаюсь на родину.
Зря она поддалась на обман. Все, с кем они с Марком встречались – по крайней мере, насколько мы можем судить, – просто обладали такой же, как у них, митохондриальной ДНК. Вообще-то Марку к тому времени уже сказали, что его К-хромосома родом из Европы (сначала это его огорчило, но позднее он обнаружил желанные африканские корни в своих митохондриях). У Болы, конечно, нет К-хромосомы, и, судя по всему, никто не потрудился взглянуть на К-хромосому ее отца, хотя это было бы любопытно: ее кожа довольно светлая. Никто не объяснил ни телеаудитории, ни Боле, ни Марку, что у остальных их генов, лежащих вне митохондрий, почти наверняка много “родин” вовсе не там, куда поместили ее создатели фильма. Если кто-нибудь проследил бы родословные других генов, столь же эмоциональное “воссоединение” могло бы произойти в любой части Африки, Европы и, скорее всего, даже Азии. Но это, конечно, лишило бы фильм драматизма.
Я не раз говорил, что использование лишь одного гена может привести к ошибкам. Зато данные, полученные от множества генов, дают нам мощный инструмент. Генные деревья для популяции и точки их ветвления отражают события прошлого. Мы можем не только установить точки ветвления, но и оценить их возраст с помощью “молекулярных часов”. И здесь ключ к разгадке, потому что схема ветвления генных деревьев, привязанная ко времени, может сказать очень о многом. Случайное спаривание (см. “Рассказ Тасманийца”) дает совсем не такой узор ветвлений, как разного рода неслучайные спаривания – каждое из них придает генному дереву особенную форму. Колебания численности популяции тоже оставляют характерный отпечаток. Таким образом, на основе нынешних паттернов распространения генов можно реконструировать прошлое, сделать выводы о размере популяций и датировать миграции. Когда популяция небольшая, дерево ветвится чаще. Увеличение популяции приводит к тому, что у деревьев отрастают длинные ветви, а точки ветвления концентрируются у основания дерева, где численность популяции еще невелика. Это свойство мы можем использовать, чтобы с помощью “молекулярных часов” определить, когда популяция увеличивалась, а когда уменьшалась, проходя сквозь “бутылочное горлышко”. Хотя, к сожалению, самые узкие “горлышки” не только стирают генетические линии, но и часто уничтожают все следы того, что было до них.
Коалесцентные генные деревья помогли разрешить давние споры о происхождении человека. Согласно “африканской теории”, все люди, живущие за пределами Африки, – потомки одной волны миграции (около 100 тыс. лет назад). Другой крайности держатся сторонники “мультирегионального” происхождения человека, считающие, что ныне живущие расы, скажем в Азии, Австралии и Европе, разделились очень давно и происходят от местных популяций Homo erectus, которые эволюционировали независимо. Оба названия сбивают с толку. Название “африканской” теории неудачно потому, что никто не спорит с тем, что все наши предки – из Африки. “Мультирегиональная” – тоже не лучшее название для теории, потому что самая древняя предковая популяция была единой. Сторонники двух этих теорий спорят о времени, когда мы покинули Африку. Так что, по-моему, лучше назвать теории так: “недавно из Африки” (“недавний исход”) и “давно из Африки” (“давний исход”). Дополнительный плюс таких названий в том, что они подчеркивают связь между ними.
Если бы все современные неафриканцы были потомками одной-единственной волны эмиграции из Африки, современные паттерны распределения генов указывали бы на недавнее прохождение небольшой африканской популяцией “бутылочного горлышка”. И точки ветвления генных деревьев тогда были бы сосредоточены вокруг даты массовой эмиграции. Если же мы независимо появились из региональных популяций H. erectus, то гены, напротив, должны указывать на то, что в каждом регионе существуют свои давно разделившиеся генетические линии. И тогда, вопреки утверждениям сторонников “недавнего исхода” о массовой эмиграции, мы почти не обнаружим точек ветвления. Что же мы видим на самом деле?
Ожидать единственного ответа на этот вопрос значило бы попасться в ту же ловушку, что и авторы фильма “Родина”. У всех генов своя история. Вполне возможно, что некоторые наши гены действительно из Африки, в то время как другие достались нам от разных популяций H. erectus. Иными словами, мы можем одновременно быть потомками и африканских эмигрантов, и региональных популяций H. erectus, потому что в любой момент в прошлом мы можем обнаружить огромное количество своих предков. Некоторые, возможно, недавно покинули Африку. Другие могли тысячи лет жить, скажем, на Яве. От первых мы могли унаследовать африканские, от вторых – яванские гены. Отдельный участок ДНК, такой как митохондриальная ДНК или К-хромосома, дает столь же скудное представление о прошлом, как отдельная фраза из учебника истории. И все же сторонники теории “недавнего исхода” часто опираются именно на данные Митохондриальной Евы. Но что будет, если “опросить” других членов “парламента” генов?
По сути, именно это сделал эволюционный биолог Алан Темплтон, предложивший теорию с удачным названием “Из Африки – снова и снова”. Темплтон использовал коалесцентную модель (подобную той, которой мы воспользовались, обсуждая гемофилию), однако применил ее не к одному гену, а к множеству. Это позволило реконструировать историю и географию генов по всему миру на отрезке в несколько сотен тысяч лет. На сегодняшний день я отдаю предпочтение Темплтону: мне кажется, он использует всю доступную информацию способом, который позволяет делать максимально обоснованные выводы. Кроме того, на каждом этапе работы он оглядывается назад, чтобы избежать злоупотребления данными.
Темплтон изучил литературу по генетике, по самым строгим критериям отобрав лишь нужное: крупные исследования в области генетики человека, в которых образцы собирали в Европе, Азии, Африке и так далее. Изучаемые гены относились к долгоживущим гаплотипам. Гаплотип – это участок генома, который либо защищен от изменений, возникающих в ходе рекомбинации (как в случае Т-хромосомы и митохондриальной ДНК), либо может оставаться неизменным достаточное для нас время (случай некоторых других, меньших участков генома). Гаплотип – это долгоживущий, легко узнаваемый участок генома. Можно представить его как “большой ген”.
Темплтон рассмотрел тринадцать гаплотипов. Для каждого он построил “генное дерево” и датировал точки ветвления с помощью “молекулярных часов”, которые были откалиброваны в основном по ископаемым. На основе датировок и географического распределения образцов он реконструировал несколько миллионов лет генетической истории нашего вида.
Главный вывод Темплтона в том, что основных волн миграции из Африки было не две, а три. В дополнение к “давнему исходу” Homo erectus около 1,7 млн лет назад (эта дата общепризнанна и основана главным образом на датировках ископаемых остатков) и недавней волне миграции, которая соответствует теории “недавнего исхода”, 840–420 тыс. лет назад случился Великий поход из Африки в Азию. Эта средняя волна миграции (“средний исход”?) подтверждается дошедшими до нас “сигналами” трех гаплотипов из тринадцати. Недавний исход подтверждается данными митохондриальной ДНК и Т-хромосом. Другие генетические “сигналы” указывают на масштабную обратную миграцию из Азии в Африку около 50 тыс. лет назад. Кроме того, митохондриальная ДНК и некоторые “небольшие” гены говорят о том, что немного позднее были и другие волны миграции: из Южной Европы в Северную, из Южной Азии в Северную, через Тихий океан в Австралию. И, наконец, согласно данным митохондриальной ДНК и археологическим находкам, около 14 тыс. лет назад произошло переселение из Северо-Восточной Азии в Северную Америку – через существовавший тогда Берингов перешеек. Вскоре после этого произошла колонизация Южной Америки – через Панамский перешеек. Кстати, говорить, что Америку открыл Христофор Колумб или Лейф Эриксон, – не что иное, как расизм. Не менее неприятно, на мой взгляд, релятивистское “уважение” к преданиям американских индейцев, которые по своему невежеству отказываются признавать, что их предки могли жить где-либо, кроме Америки.
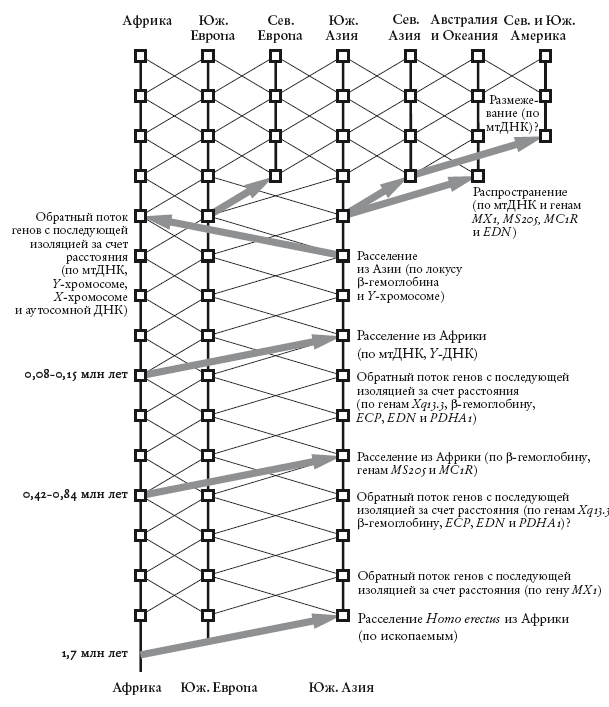
Из Африки – снова и снова. Схема основных миграций человека, построенная А. Темплтоном на основе изучения 13 гаплотипов. Вертикальные линии соответствуют наследованию, диагональные отображают генетический перенос. Стрелками показаны миграции, подтвержденные данными генетических исследований. TEMPLETON [284].
В промежутках между тремя темплтоновскими волнами, как показывают генетические данные, происходил непрерывный перенос генов между Африкой, Южной Европой и Южной Азией. Согласно данным Темплтона, крупные и мелкие волны миграции сопровождались скрещиванием с местным населением, а не полным истреблением той или иной стороны – хотя, возможно, и такое бывало. Это очень важно для понимания нашей родословной.
Наш рассказ, как и исследование Темплтона, посвящен людям и их генам. Однако родословные есть не только у человека. Все виды наследуют генетический материал, и у всех видов с половым размножением есть свои Адам и Ева. Методы изучения недавней истории человечества применимы и к остальным формам жизни. Так, ДНК гепарда “рассказала”, что 12 тыс. лет назад популяция прошла сквозь “бутылочное горлышко”. Эта информация важна для специалистов по охране кошачьих. В ДНК кукурузы сохранились несомненные свидетельства ее одомашнивания, которое произошло в Мексике 9 тыс. лет назад. Паттерны ветвления у деревьев штаммов ВИЧ могут использовать эпидемиологи и врачи, чтобы понять принципы распространения вируса и научиться его сдерживать. Гены и генные деревья проливают свет на историю флоры и фауны Европы: они рассказывают о массовых миграциях, вызванных ледниковыми периодами, которые постепенно оттесняли виды умеренного климата в южноевропейские убежища (рефугиумы) и оставляли арктические виды в изолированных горных областях. Эти и другие события можно проследить, изучая данные о распределении ДНК по земному шару. Эти данные составляют исторический справочник, который нам еще предстоит прочесть.
Мы видели, что каждый ген может рассказать свою историю; совместив эти истории, можно реконструировать события прошлого – недавнего и далекого. Древнейшие ПОП наших генов могут относиться к тому времени, когда мы еще не были людьми.
Происходит это, в частности, тогда, когда естественный отбор благоприятствует разнообразию популяции в ее интересах.
Предположим, есть две группы крови, A и B, и каждая обеспечивает иммунитет к разным заболеваниям. Каждая группа крови восприимчива к тому заболеванию, против которого у другой группы крови есть иммунитет. Заболевание распространяется в том случае, когда в популяции широко распространены носители восприимчивой к ней группы крови: в этом случае может начаться эпидемия. Поэтому, если в популяции преобладают люди, например, с группой крови B, заболевание легко перерастет в эпидемию. В результате люди с группой крови B будут умирать, пока их не станет совсем мало, а людей с группой крови А – соответственно, много. Если существует два типа какого-либо признака, более редкому из которых благоприятствует отбор просто в силу его редкости, это и есть рецепт полиморфизма: поддержание разнообразия ради разнообразия. Система групп крови АВ о – всем известный полиморфизм, который, возможно, сохраняется именно по указанным причинам.
Некоторые полиморфизмы могут быть довольно стабильными – настолько стабильными, что они передаются от предкового вида дочернему. Удивительно, но наш полиморфизм АВ о есть и у шимпанзе. Может статься, что мы с шимпанзе “изобрели” его независимо по одной и той же причине. Но, скорее всего, мы унаследовали его от общего предка и сохранили в течение 6 млн лет независимого развития, поскольку все это время соответствующие заболевания никуда не исчезали. Это межвидовой полиморфизм, и он наблюдается у видов куда более далеких, чем мы и шимпанзе.
Потрясающе: в отношении некоторых генов мы ближе к шимпанзе, чем к людям. Я, например, могу оказаться ближе к отдельным шимпанзе, чем к вам (или вашему шимпанзе). Человек как вид и как особь – лишь временный сосуд с генами. Индивид – остановка на перекрестке путей, которыми гены идут сквозь историю. Именно так можно выразить основную идею моей книги “Эгоистичный ген”. Там я писал: “После того как мы выполнили свою задачу, нас выбрасывают. Но гены – выходцы из геологического времени, они здесь навеки”. Однажды я выразил эту идею в стихах:
Ген-странник мне сказал:
“Тел много я видал.
Не будь таким беспечным:
Я существую вечно,
А ты – всего лишь тело
Для выживанья гена”.
А в ответе тела я спародировал “Песнь датчанок” Киплинга:
Что тело – если ты покидаешь его,
Проведя в нем короткий миг.
И уходишь туда, где тебя одного
Ждет старый слепой часовщик…
Мы назначили рандеву № о несколько десятков тысяч лет назад, максимум – несколько сотен тысяч лет назад. Туда мы еще не дошли. Следующее рандеву, где мы встретимся с шимпанзе, случилось на много миллионов лет раньше, а большинство рандеву вообще в сотнях миллионов лет от нас. Чтобы вовремя завершить паломничество, нужно поспешить. Мы, не останавливаясь, минуем около тридцати ледниковых периодов, типичных для последних 3 млн лет, и минуем эпохи перемен, когда пересыхало и снова наполнялось Средиземное море (4,5–6 млн лет назад). Когда нам захочется передохнуть, я воспользуюсь правом останавливаться в промежуточных точках, где мертвые ископаемые будут рассказывать нам истории. “Призраки” других пилигримов, с которыми мы встретимся, и их истории помогут нам удовлетворить законный интерес к собственным предкам.
Назад: Рассказ Тасманийца[2]
Дальше: Архаичный Homo sapiens

