Книга: Рождение сложности. Эволюционная биология сегодня: неожиданные открытия и новые вопросы
Назад: Вирусы и мобильные генетические элементы
Дальше: Что почитать на эту тему в Интернете
Эпигенетическое наследование
Недавно открыто еще несколько молекулярных механизмов, которые в принципе могут служить для передачи по наследству приобретенных признаков. Эти механизмы не связаны напрямую с изменениями самого «текста», записанного в структуре молекул ДНК, то есть с мутациями. Поэтому такую наследственность называют «эпигенетической», или «надгенетической».
Один из эпигенетических механизмов — метилирование ДНК. Оказалось, что в процессе жизнедеятельности к молекулам ДНК в клетках, в том числе и в половых, специальные ферменты «пришивают» метильные группы (-CH3). Причем к одним генам метильных групп пришивают больше, к другим — меньше. Распределение метильных групп по генам зависит от того, насколько активно тот или иной ген используется. Активные гены метилируются слабо, неактивные — сильно. Получается совсем как с упражнением и неупражнением органов, которое Ламарк считал причиной наследственных изменений. Поскольку «рисунок метилирования» может передаваться по наследству и поскольку он, в свою очередь, влияет на активность генов у потомства, легко заметить, что здесь может работать совершенно ламарковский механизм наследования: «натренированные» предками гены будут и у потомства работать активнее, чем «ослабевшие» от долгого неиспользования.
Однако и в данном случае природа, судя по всему, не заинтересовалась возможностью наладить наследование «по Ламарку». Наибольшего развития система метилирования ДНК в половых клетках достигла у двух групп высших многоклеточных, венчающих эволюционное древо, соответственно, животных и растений — а именно у плацентарных млекопитающих и покрытосеменных (цветковых) растений. В обоих случаях избирательное метилирование ДНК в половых клетках, так называемый «геномный импринтинг», служит не для передачи по наследству приобретенных признаков, а для регуляции взаимоотношений между зародышем и материнским организмом.
—————Геномный импринтинг — распространенное у млекопитающих и цветковых растений явление, состоящее в том, что некоторые гены в половых клетках родителей особым образом «метятся». К цитозиновым основаниям прикрепляется метильная группа. «Помеченный» ген у потомства просто-напросто не работает. Некоторые гены отключаются в сперматозоидах, другие в яйцеклетках. Если гены отключены в сперматозоидах, то соответствующие признаки потомство наследует только от матери. Если отключены гены в яйцеклетке, то наследуются отцовские признаки. В половых клетках потомства старые метки удаляются и заменяются новыми. В результате у внуков могут проявиться признаки деда или бабки, которые не были выражены у родителей. Импринтинг — это пример так называемой «эпигенетической» наследственности, то есть наследственных свойств, не связанных с изменением последовательности нуклеотидов в молекулах ДНК, генетического текста.Именно благодаря импринтингу при скрещивании разных пород или видов млекопитающих оказывается небезразлично направление гибридизации, то есть кто из двух родителей будет принадлежать к какому виду. Например, при скрещивании осла с кобылой получаются мулы, а при скрещивании жеребца с ослицей — лошаки. Вопреки классическим законам генетики важным для потомства оказывается не только то, какие гены они получили, но и от кого — от отца или матери.Зачем нужен геномный импринтинг, почему он появился? Для объяснения этого существует две гипотезы. Первая — общепринятая — состоит в том, что импринтинг развился вследствие различия интересов полов. У млекопитающих между самкой и ее детенышем во время внутриутробного развития складываются отчасти антагонистические отношения. Говоря упрощенно, эмбрион старается высосать из матери побольше соков, а мать старается сохранить силы и здоровье, чтобы в будущем иметь возможность родить и других детей. Самец в этом конфликте в общем случае — на стороне детеныша. Других-то детей самка еще неизвестно от кого родит, а этот — свой. Поэтому самцы отключают в своих сперматозоидах те гены, которые способствуют защите матери от чрезмерных притязаний эмбриона, а матери, напротив, отключают в своих яйцеклетках те гены, которые могут эти притязания усилить. Действительно, большинство генов млекопитающих, подвергающихся родительскому импринтингу, так или иначе связаны с внутриутробным развитием, строением плаценты и т. п.Из-за этого у млекопитающих (в отличие от многих других животных, таких как насекомые или ящерицы) принципиально невозможно развитие зародыша на основе только материнских или только отцовских генов.Можно взять только что оплодотворенную яйцеклетку какого- нибудь другого животного (не млекопитающего, а, например, лягушки), в которой ядра яйцеклетки и сперматозоида еще не успели слиться, удалить из нее отцовское ядро и заменить его ядром из другой яйцеклетки. Два «материнских» ядра сольются, и из такой яйцеклетки в благоприятных условиях может развиться нормальный организм, все гены которого — материнские. Можно искусственно сделать яйцеклетку с двумя отцовскими ядрами, и из нее тоже может вырасти жизнеспособное животное. Однако с млекопитающими этот номер не пройдет. Без отцовских генов у эмбриона не разовьется плацента, а без материнских плацента разовьется даже лучше, чем надо, но не будет нормально развиваться сам эмбрион.Дополнительная гипотеза, более симпатичная с этической точки зрения, была высказана совсем недавно. Согласно этой гипотезе основной смысл геномного импринтинга — достижение лучшей совместимости между матерью и плодом, то есть на первое место ставятся не антагонистические взаимоотношения матери и эмбриона, а кооперативные. Если часть отцовских генов будет выключена, то у эмбриона будут работать только материнские копии этих генов, и эмбрион, таким образом, будет по своим физиологическим и биохимическим свойствам больше похож на мать, и им легче будет приспособиться друг к другу. Эта гипотеза предполагает, что в ходе родительского импринтинга должно отключаться больше отцовских генов, чем материнских, и факты это подтверждают.—————
Метилирование и деметилирование генов играют важную роль в процессе индивидуального развития многоклеточного организма, где они служат для контроля работы генов в развивающихся органах и тканях. Кроме того, при помощи метилирования клетки борются с чрезмерной активностью мобильных генетических элементов. Избирательное метилирование МГЭ снижает частоту их «прыжков». Особенно тщательно многоклеточные организмы «следят» за активностью МГЭ при созревании сперматозоидов (см. главу «На подступах к неведомому»).
Очень важно, что метилирование дает клетке возможность попутно контролировать и частоту мутирования тех или иных генов. Как уже отмечалось в главе «Управляемые мутации», метилирование цитозина (Ц) резко повышает вероятность мутации, в результате которой цитозин заменяется тимином (Т). Метилирование ДНК активно контролируется клеткой, таким образом, в клетке реально существует механизм, позволяющий целенаправленно регулировать вероятность мутирования определенных участков генома.
—————Взаимное включение и выключение генов. Еще один возможный вариант эпигенетического наследования приобретенных признаков основан на системах взаимной активации и инактивации генов. Допустим, ген А производит белок, одно из действий которого состоит в блокировании работы гена Б, а ген Б, в свою очередь, кодирует другой белок, способный «выключать» ген А. Такая система может находиться в одном из двух состояний: либо ген А работает, и тогда ген Б выключен, либо наоборот. Допустим, что переход системы из одного состояния в другое может происходить только в результате какого-то особенного внешнего воздействия, и случается такое редко. То состояние, в котором находится эта двухгенная система в клетках матери, будет через яйцеклетку передаваться ее потомству (поскольку сперматозоид содержит пренебрежимо малое количество белков). Если же в течение жизни матери система переключится в другое состояние, то этот приобретенный признак передастся потомству, родившемуся после «переключения». Опять получается наследование по Ламарку. Точнее, возможность такого наследования. И опять, как и в предыдущих случаях, живые организмы, похоже, не очень-то торопятся воспользоваться этой замечательной возможностью.—————
Мы видим, что «ламарковское» наследование приобретенных признаков вполне осуществимо технически. Имеется целый ряд молекулярных механизмов, способных обеспечить целенаправленную передачу потомству наследуемой информации о приобретенных признаках. Тот факт, что живые организмы редко используют эти возможности, говорит о том, что наследование «по Ламарку» им просто не выгодно.
—————Негенетическая «память поколений». Передача информации от родителей к потомкам может осуществляться тремя основными путями, два из которых общеизвестны: это генетическая наследственность, свойственная всем без исключения живым организмам, и обучение, характерное только для животных со сложной нервной системой.Третий путь менее известен и гораздо хуже изучен, однако и он, судя по всему, играет важную роль в жизни многих организмов. Это так называемые «родительские эффекты» — внегенетические изменения у потомства, обусловленные условиями жизни и заботой родителей. Простейший пример — самка, плохо питавшаяся в течение своей жизни, откладывает яйца с меньшим количеством питательных веществ, из которых развивается — даже при «хороших» генах — сравнительно чахлое потомство. Чем не наследование «приобретенного признака»?Более сложные варианты могут включать различные эпигенетические изменения наследственного материала (ДНК), о которых мы говорили выше, в том числе геномный импринтинг, который представляет собой не что иное, как целенаправленное манипулирование наследственными свойствами потомства. Кроме того, известно, что эмбриональное развитие животных, особенно на начальных этапах, во многом зависит от разнообразных молекул (в том числе матричных РНК), поступающих в яйцеклетку из материнского организма (см. раздел «Нужны ли эмбрионам гены» в главе 5). Ясно, что условия жизни матери в принципе могут влиять на количество и состав этих веществ и, следовательно, на развитие зародыша.Изучать родительские эффекты легче всего у тех животных, в жизненном цикле которых присутствует партеногенетическое размножение (развитие потомства из неоплодотворенныхяиц) — как, например, у рачков-дафний. В этом случае геномы матери и ее дочерей идентичны и легче отличить «материнские» эффекты от генетических.Недавно российским ученым удалось показать методом моделирования, что материнский эффект — негенетическая передача от матери к потомству информации о длине светового дня и обилии пищи — играет важную роль в сезонных изменениях численности и поведения дафний и делает популяцию более устойчивой. Наличие материнского эффекта у дафний, вначале предсказанное теоретически, недавно получило экспериментальные подтверждения.Еще в 1996 году А. А. Умнов и В. Р. Алексеев разработали имитационную модель для проверки гипотезы о существовании у ветвистоусых рачков материнского эффекта. В модели предполагалось, что дафнии передают потомству информацию о трофических условиях (то есть о том, насколько хорошо питалась мать). Позже эта гипотеза была подтверждена экспериментально. Выяснилось, что дафнии передают потомству информацию не только о своем питании, но и об изменениях длины светового дня. Вследствие этого особи, имеющие одинаковый размер и возраст, могут по-разному реагировать на одни и те же пищевые и температурные условия только потому, что их матери имели разную жизненную историю.
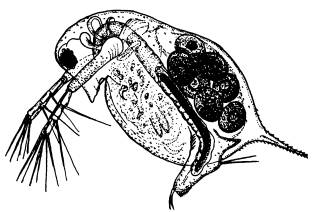
Ветвистоусый рачок Daphnia longispina — массовый представитель зоопланктона озер и луж умеренной зоны. На фотографии видны зреющие яйца в выводковой камере.
В новой статье В. Р. Алексеева и Т. И. Казанцевой, опубликованной в 2007 году в «Журнале общей биологии», оценивается возможное влияние материнского эффекта на колебания численности популяций дафний, на время перехода от партеногенетического (однополого) размножения к двуполому, на количество зимующих яиц и другие параметры популяции.В качестве «образца» для моделирования использовалась хорошо изученная популяция рачков Daphnia longispina из озера Красненького в Псковской области. Жизненный цикл этих рачков довольно сложен. Весной из зимующих яиц выходит первое поколение самок, которые начинают быстро размножаться путем партеногенеза, производя на свет только самок. В начале лета численность микроскопических водорослей — основной пищи дафний — снижается, и дафнии начинают производить на свет самцов и переходят к двуполому размножению. Образуются покоящиеся оплодотворенные яйца, и наступает «летняя диапауза», длящаяся не более месяца. Затем из яиц выходит новое поколение самок, размножающихся партеногенетически. В конце лета и осенью в популяции снова появляются самцы, начинается двуполое размножение и откладываются зимующие яйца, покрытые плотной оболочкой, позволяющей переносить неблагоприятные условия.Переход дафний от однополого размножения к двуполому регулируется несколькими факторами — температурой, обилием пищи, длиной светового дня. Информация о динамике этих факторов в прошлом и настоящем доходит до дафний двумя путями: из их личного жизненного опыта, а также от матери благодаря «материнскому эффекту».Разработанная авторами модель имитирует жизненный путь каждой отдельной особи. Вероятность перехода особи от однополого размножения к двуполому зависит в модели как от сиюминутного состояния факторов среды (температуры, длины светового дня, количества пищи), так и от индивидуального «потенциала роста» особи, который, в свою очередь, определяется условиями жизни и «потенциалом» ее матери. Потенциал роста определяет максимально возможную скорость роста данной особи. Чем медленнее рост, тем выше вероятность перехода к двуполому размножению.Таким образом, переход к двуполому размножению зависит не только оттого, как питалась данная особь и каковы условия среды в данный момент, но и оттого, как питались ее мать и более далекие предки. Тем самым задается «материнский эффект» — негенетическая передача информации от матери к потомкам. Авторам удалось показать, что при определенных входных параметрах их модель очень точно воспроизводит реальную динамику численности самцов и самок, партеногенетических и покоящихся яиц, наблюдаемую в реальной популяции. Надо сказать, что эта реальная динамика достаточно сложна: например, в течение одного лета наблюдается несколько (обычно 5) пиков численности дафний.Точное соответствие модельной и реальной популяционной динамики дало основания полагать, что модель действительно учитывает все основные факторы и причинно-следственные связи, определяющие эту динамику. Теперь модель можно было использовать для проверки гипотез о роли материнского эффекта.Изъятие из модели материнского эффекта привело к тому, что модельная динамика стала резко отличаться от реальной. Ни один из вариантов модели с отключенным материнским эффектом не позволил воспроизвести динамику численности и формирование банка покоящихся яиц, необходимых для устойчивого многолетнего развития популяции.Например, если потенциал роста был задан высоким, покоящиеся яйца практически не образовывались, выживание популяции в зимний период было возможно только благодаря небольшому количеству перезимовавших самок и в долгосрочной перспективе популяция становилась крайне уязвимой. На этом основании авторы сделали вывод о существенной роли материнского эффекта в формировании популяционной динамики и сезонных адаптаций у дафний.(Источник: В. Р. Алексеев, Т. И. Казанцева. Использование индивидуально-ориентированной модели для изучения роли материнского эффекта в смене типов размножения у Cladocera // Журнал общей биологии. 2007. Т. 68. С. 231–240.)—————