Книга: Рождение сложности. Эволюционная биология сегодня: неожиданные открытия и новые вопросы
Назад: Видообразование на разных островах идет параллельными путями
Дальше: Закономерная повторяемость в неповторимом живом многообразии
Что заставляет эволюцию повторяться?
Для начала отметим первое, что бросается в глаза. Сходство параллельно развивающихся групп обычно тем больше, чем ближе их родство. Очень далекие друг от друга группы могут выработать лишь самое поверхностное сходство (например, рыбы и дельфины). Наоборот, в Большом и Малом Арале или на крошечных островках архипелага Тристан-да-Кунья сходство вновь образующихся форм было очень велико, поскольку они происходили от одних и тех же исходных видов. По-видимому, само строение организма во многом предопределяет возможные пути его эволюции. Для каждого организма существует весьма ограниченное число «разрешенных» путей эволюционных изменений — и любые попытки уклониться от этих путей безжалостно отсекаются отбором. У близкородственных видов, индивидуальное развитие которых идет по сходной «программе», набор этих разрешенных путей тоже сходен, что и приводит к параллелизмам.
Во-вторых, большую роль играет сходство структуры основных типов сообществ, или экосистем. Набор экологических «ролей» (ниш) в разных сообществах в целом сходен. Например, если это степь, прерия или саванна, то там должна быть, во-первых, трава, во-вторых — крупные травоядные звери (копытные или их аналоги), в третьих — хищники, охотящиеся на этих копытных, в четвертых — жуки-навозники и другие беспозвоночные, перерабатывающие навоз травоядных, и т. д. Все эти ниши обязательно должны быть кем-то заполнены, иначе весь комплекс развалится. Попробуйте лишить экосистему одного из блоков, например жуков-навозников, — немедленно начнется экологический кризис. Нечто подобное наблюдалось в Австралии, где завезенные европейскими фермерами овцы стали быстро приводить пастбища в негодность из-за отсутствия в местной фауне эффективных переработчиков навоза. Пришлось срочно завозить жуков-навозников с «Большой земли». У фермеров не было времени ждать, пока эволюция справится с проблемой в своей обычной неторопливой манере. А подождали бы пару сотен тысячелетий — и почти наверняка на основе местных австралийских жуков развились бы «свои» эффективные навозники. И они, скорее всего, по многим признакам были бы похожи на своих коллег с других континентов. Обитание в сходных условиях и нацеленность на сходный образ жизни приводят к тому, что организмы приобретают сходные черты строения.
Кроме того, в последнее время появляется все больше свидетельств, что «случайные и ненаправленные» наследственные мутации, которые, согласно господствовавшим до недавнего времени представлениям, являются единственным источником материала для отбора, в действительности часто оказываются не такими уж случайными. Об этом мы поговорим подробно в главе «Управляемые мутации». Конечно, природе постоянно приходится действовать «методом случайного тыка», но она всячески старается этот метод оптимизировать.
Наконец, многие параллелизмы могут иметь довольно простые генетические объяснения. Индивидуальное развитие животных и растений контролируется сравнительно небольшим числом ключевых генов-регуляторов. Если, например, в двух эволюционных линиях независимо «сломается» какой-то регуляторный ген, в этом едва ли можно усмотреть что-то удивительное или невероятное. Однако внешнее проявление этой ошибки может показаться в высшей степени удивительным: ведь мы увидим весьма сложные и, главное, одинаковые изменения в двух разных группах организмов.
Один из ярких примеров такого рода дают поясничные ребра, которые на ранних этапах эволюции млекопитающих многократно исчезали и снова появлялись в разных эволюционных линиях. Недавно китайские палеонтологи нашли очередного доисторического зверька, строение которого проливает новый свет на это явление. Данная находка, помимо прочего, представляет собой прекрасный пример классической «переходной формы».
Надо сказать, что находки древних мезозойских млекопитающих в последнее время необычайно участились. Еще пару десятилетий назад считалось, что мезозойские млекопитающие были малочисленной, второстепенной группой мелких крысоподобных зверьков, влачивших жалкое существование в тени господствовавших в то время динозавров. Благодаря находкам последних лет, значительная часть которых сделана китайскими палеонтологами, стало известно, что древние звери были весьма многочисленны, разнообразны и могли достигать размеров лисы или барсука.
—————Новая палеонтологическая находка проливает свет на раннюю эволюцию млекопитающих. Китайские палеонтологи в очередной раз удивили научный мир необычной находкой. На этот раз они откопали очень хорошо сохранившийся скелет неизвестного ранее примитивного млекопитающего из группы триконодонтов, жившего около 125 млн лет назад (в раннемеловую эпоху).Новооткрытый мезозойский зверек Yanoconodon allini был как раз таким, каким «положено» быть мезозойскому зверю — маленьким и невзрачным. Интересен он прежде всего тем, что у него очень хорошо сохранились косточки среднего уха (молоточек и наковальня), причем сохранились они не по отдельности, а в своем естественном положении, сочлененные с другими костями.Строение этих косточек является одним из главных отличительных признаков млекопитающих. У предков зверей — зверозубых рептилий — была только одна слуховая косточка (стремечко), а молоточек и наковальня составляли единое целое с нижней челюстью. То же самое наблюдается и у самых примитивных древних млекопитающих. У современных млекопитающих в ходе эмбрионального развития эти косточки сначала, как и у далеких предков, прилегают к нижней челюсти, а затем отделяются от нее. Отделение происходит в два этапа: сначала слуховые косточки отделяются от боковой поверхности нижней челюсти, но остаются связаны с ней спереди посредством меккелева хряща (этот хрящ, кстати, представляет собой остаток первичной, изначальной нижней челюсти позвоночных; у акул вся нижняя челюсть образована меккелевым хрящом). На втором этапе исчезает и это переднее соединение: меккелев хрящ у взрослых млекопитающих рассасывается.На основе этих фактов более 30 лет назад была выдвинута гипотеза, согласно которой в эволюции среднего уха млекопитающих имелась промежуточная стадия — примерно такая же, как в эмбриональном развитии современных зверей. Яноконодон блестяще подтвердил эту догадку. Именно поэтому первооткрыватели присвоили ему видовое название allini — в честь Эдгара Аллина (Edgar Allin), автора подтвердившейся гипотезы. Молоточек и наковальня у яноконодона уже отделены от нижней челюсти с боков, но еще прикреплены к ней спереди посредством окостеневшего меккелева хряща.Строение скелета яноконодона еще раз подтвердило, что в ранней эволюции млекопитающих многие прогрессивные признаки появлялись параллельно и независимо в разных линиях. Например, у яноконодона имеются ребра на поясничных позвонках (это примитивный признак), тогда как у некоторых близких форм поясничные ребра отсутствуют. Сравнительно-анатомический и филогенетический (эволюционный) анализ мезозойских млекопитающих показывает, что поясничные ребра многократно исчезали и вновь появлялись в разных эволюционных линиях.По мнению исследователей, в случае с поясничными ребрами, как и с другими признаками, затрагивающими дифференцировку отделов позвоночника, многочисленные параллелизмы могут объясняться тем, что эти признаки контролируются очень небольшим количеством генов. Речь идет о знаменитых Hox-генах, многофункциональных регуляторах развития, которые задают общий план строения, симметрию и дифференцировку отделов тела у многоклеточных животных. Например, если у мыши отключить ген Hox10 (который, правда, присутствует в геноме в трех копиях, и отключить надо все три), то на поясничном отделе позвоночника развиваются ребра. Скорее всего, многократное появление и исчезновение поясничных ребер у древних млекопитающих объясняется простотой «кодирования» этого признака, то есть тем, что для его изменения достаточно всего одной или немногих мутаций.
Yanoconodon allini.
Сравнительно простые, эффективные и «умные» системы регуляции индивидуального развития, сложившиеся в ходе эволюции у животных (в том числе система Hox-генов), обладают важным побочным свойством: их случайные изменения (например, в результате мутаций) с довольно высокой вероятностью могут породить не только нежизнеспособных монстров, но и что-то вполне осмысленное и даже удачное. В этом, возможно, состоит одна из причин ускорения прогрессивной эволюции у высших животных.(Источник: Zhe-Xi Luo, Peiji Chen, Gang Li, Meng Chen. A new eutriconodont mammal and evolutionary development in early mammals // Nature. 2007. V. 446. P. 288–293.)—————

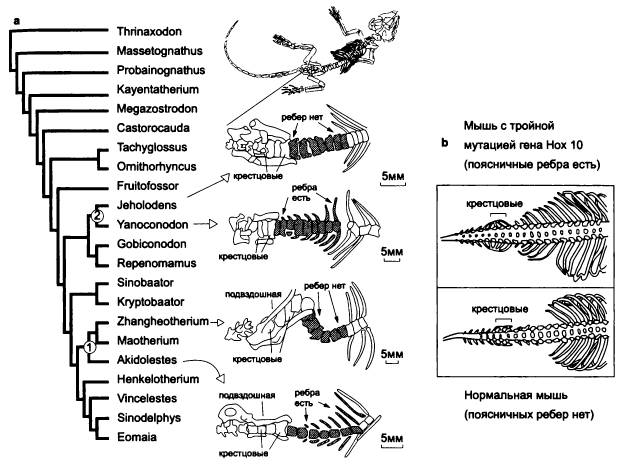
Параллельная эволюция признака «наличие/отсутствие поясничных ребер» у мезозойских млекопитающих.
Генетические основы параллельной эволюции изучены пока слабо. Сходные изменения морфологии или биохимии организмов могут быть обусловлены как одинаковыми, так и совершенно разными наборами мутаций. Какой из этих двух вариантов встречается чаще, до сих пор не известно, и число детально исследованных случаев невелико.
В главе «Рождение сложности» мы говорили о том, как бактерии приспосабливаются к антибиотикам. Такое приспособление происходит у разных бактерий параллельно и, по-видимому, может обеспечиваться одним и тем же комплексом мутаций, приобретаемых в определенной последовательности.
Однако это крайне простой и редкий случай, когда между признаком (устойчивость к антибиотику) и геном (в данном случае это ген фермента бета-лактамазы) наблюдается соотношение один к одному. Гораздо чаще, особенно у организмов, более сложных, чем бактерии, каждый ген влияет на множество признаков, а каждый признак определяется множеством генов. К тому же связи между генами и признаками опосредуются многочисленными внешними и внутренними факторами в ходе индивидуального развития. Все это делает задачу «расшифровки» генетических основ эволюционных преобразований чрезвычайно сложной.
—————Тонкая подстройка многофункционального гена может приводить к параллельному появлению новых признаков. Группе исследователей из США удалось расшифровать генетический механизм параллельной эволюции пятен на крыльях у самцов мушек-дрозофил. Эти пятна имеются у одних видов рода Drosophila и отсутствуют у других. В процессе эволюции пятна появлялись и утрачивались неоднократно, хотя выглядят они у всех видов примерно одинаково.Исследователи показали, что наличие или отсутствие пятен определяется многофункциональным геном yellow («желтый»), от которого зависит окраска не только крыльев, но и прочих частей тела. При этом пятнистость крыльев определяется не «основным» (кодирующим) участком гена, в котором записано строение соответствующего белка. Как выяснилось, все дело в том, в каких частях развивающегося крыла происходит экспрессия (работа) гена yellow на стадии куколки. У видов без пятен слабая экспрессия наблюдается равномерно по всему крылу, резко усилена на концах крыльев (как раз там, где у взрослой мухи будут темные пятна).
В ходе эволюции пятна на крыльях дрозофилы несколько раз утрачивались и появлялись — в зависимости от характера экспрессии многофункционального гена yellow.
Характер экспрессии гена yellow определяется двумя некодирующими регуляторными участками гена. Пятна на крыльях у дрозофил иногда появлялись благодаря модификации первого из этих участков, иногда — второго. Таким образом, в данном случае независимое появление одного и того же признака (пятен на крыльях) было обеспечено разными генетическими изменениями, затронувшими, правда, один и тот же ген.Исследование показало, что тонкая «подстройка» регуляторных участков многофункциональных генов может приводить к появлению новых морфологических признаков у таких сложных организмов, как дрозофила.(Источник: Benjamin Prudhomme, Nicolas Gompel, Antonis Rokas, Victoria A. Kassner, Thomas M. Williams, Shu-Dan Yeh, John R. True, Sean B. Carroll. Repeated morphological evolution through cis-regulatory changes in a pleiotropicgene // Nature. 2006. V. 440. P. 1050–1053.)—————
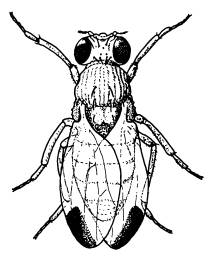
По-видимому, именно изменения регуляторных участков генов играют основную роль в эволюции формы и строения сложных организмов. Большинство генов у таких организмов — многофункциональные. Изменение кодирующей части многофункционального гена может вызвать целый каскад разнообразных малопредсказуемых последствий. Гораздо удобнее и безопаснее чуть-чуть «подкрутить» один из многочисленных регуляторов, влияющий не на все функции данного гена сразу, а лишь на одну или несколько.
Важнейший для всей человеческой цивилизации пример параллельной эволюции дают культурные злаки. Ведь все множество одомашненных видов и сортов злаков, от кукурузы до пшеницы, должны были обрести совершенно определенный набор хозяйственных признаков. Это скороспелость, высокая продуктивность, неопадающие семена, голозерность и др. Скороспелость, то есть превращение многолетников или двухлетников в однолетники, необходима, так как урожай желательно иметь каждый год, а не через два года на третий. Неопадающие семена и неломкий колос нужны для предотвращения опадения на землю спелых семян до сбора их земледельцем. Голозерность сильно облегчает и ускоряет процесс обмолота. И все эти признаки культурные растения приобрели в результате неосознанного искусственного отбора независимо друг от друга. Такая параллельная эволюция, по-видимому, сильно облегчалась тем, что формирование многих хозяйственных признаков контролируется единичными регуляторными генами. И если этот единственный регуляторный ген отключить или изменить его работу мутацией, то сразу получится нужный признак. По-видимому, именно таким способом сформировались у многих злаков неопадающие семена и неломкие стебли. Для появления подобного признака часто бывает достаточно «испортить» всего один регуляторный ген из семейства MADS-генов. MADS-гены — это гены растений, аналогичные по функциям Hox-генам животных. Эти гены у культурных растений имеют очень большое сходство, поэтому легко можно ожидать и появления сходных мутаций у разных растений.
Назад: Видообразование на разных островах идет параллельными путями
Дальше: Закономерная повторяемость в неповторимом живом многообразии

