Книга: Рождение сложности. Эволюционная биология сегодня: неожиданные открытия и новые вопросы
Назад: Пути эволюции предопределены на молекулярном уровне
Дальше: «Несократимая сложность»
Приключения Protozoon (модель возникновения сложного организма из простого)
Попробуем понять, как же все-таки в ходе эволюции сложное может рождаться из простого. Для этого я хочу предложить вниманию читателей забавную мысленную модель.
Героем нашего повествования будет вымышленное, но довольно правдоподобное одноклеточное существо Protozoon, имеющее вполне обычный жизненный цикл, всего один «орган» — глазок — и всего один внутренний регуляторный фактор (сигнальное вещество X) с двумя эффектами. Это незамысловатое простейшее, как мы увидим, в результате одной- единственной случайной мутации автоматически может дать начало целому вееру сложных многоклеточных форм со своими онтогенезами (способами индивидуального развития), жизненными циклами, разделением тела на вегетативную и генеративную части («сому» — тело и «герму» — половые клетки).
Итак, прошу познакомиться: Protozoon, диплоидный одноклеточный эукариотический организм с простым жизненным циклом. В благоприятных условиях у зиготы образуется глазок. Глазок стимулирует выработку сигнального вещества X.

Как у большинства реальных одноклеточных эукариот, клетка нашего Protozoon может делиться двумя способами: митозом (получаются две диплоидные дочерние клетки) и мейозом (получаются четыре гаплоидные половые клетки — гаметы, снабженные жгутиками и потому подвижные).
Сигнальное вещество X имеет два эффекта. В концентрации 1 оно подавляет процесс образования глазков, а в концентрации 4 и выше приводит к тому, что созревшая клетка будет делиться митозом (при меньшей концентрации — мейозом). Вещество X способно с определенной скоростью проникать сквозь мембрану (оболочку) клетки в обе стороны.
—————Митоз — деление эукариотической клетки, в результате которого из одной родительской клетки получаются две дочерние с таким же числом хромосом, как и у родительской. Например, митоз диплоидной клетки приводит к формированию двух диплоидных клеток.Мейоз — «редукционное деление» эукариотической клетки, в результате которого число хромосом сокращается вдвое. Из диплоидной родительской клетки (с двойным набором хромосом) получаются четыре гаплоидных клетки с одинарным набором хромосом. У животных таким путем образуются половые клетки — яйцеклетки и сперматозоиды.—————
Глазок к тому же придает клетке полярность. Это проявляется в том, что, когда клетка делится путем митоза, плоскость деления всегда располагается перпендикулярно тому диаметру клетки, на котором находится глазок. В результате глазок «достается» одной из дочерних клеток, а другая поначалу не имеет глазка. Будем считать, что ориентация плоскости деления задается глазком при помощи выделяемого им вещества X. Концентрация этого вещества максимальна возле глазка и постепенно снижается по мере удаления от него. Иными словами, существует «градиент концентрации» вещества X, и плоскость деления всегда перпендикулярна направлению этого градиента.
При неблагоприятных условиях зигота не образует глазка, вещество X не вырабатывается, и поэтому, если ей удастся дорасти до зрелости, она делится только мейозом и образует четыре подвижные гаметы.
Посмотрим теперь, в чем смысл этого нехитрого регуляторного контура и как складывается жизнь Protozoon в естественных условиях. Если все хорошо и пищи много, все клетки в сообществе Protozoon образуют глазок и делятся митозом. По мере роста плотности популяции увеличивается концентрация X в воде. Это вещество, как мы знаем, проходит через мембрану. Поэтому в конце концов в молодых клетках, еще не успевших образовать глазок, концентрация вещества X (впитанного из окружающей среды) достигнет значения 1. Тогда у них не образуется глазок, и они будут делиться мейозом, гаметы расплывутся, популяция разредится, и произойдет расселение. Очень несложная, вполне правдоподобная и устойчивая система.
Но вот у одной особи возникла мутация: утрачена способность клеток расходиться после митоза. Условия благоприятны. Что теперь будет? Сразу, без всякого отбора и дополнительных мутаций, появляется множество разных многоклеточных организмов со своими онтогенезами и жизненными циклами.

Это самый простой из возникших вариантов. Двуклеточный организм, у которого передняя клетка — вегетативная («сома») с глазком, а задняя — генеративная («герма»). Задняя клетка, созревая, производит гаметы и сразу же заменяется новой, образовавшейся в результате деления головной клетки.
В чем секрет? Оказывается, все дело в сигнальном веществе X. Как только у клетки появляется глазок, концентрация вещества X в ней почти мгновенно достигает максимального уровня 10. Избыточные количества вещества X интенсивно откачиваются из клетки и… проникают в соседнюю, «прилипшую» сзади вторую клетку. И там устанавливается постоянная концентрация X, равная 5. Раньше, когда клетки расходились сразу после деления, подобных ситуаций не возникало. Только теперь, когда клетки остались слипшимися, стало ясно, что проницаемость мембраны для вещества X такова, что в клетке, примыкающей к своей «соседке» с глазком, устанавливается половинная концентрация вещества X. Если к клетке с концентрацией 5 «прилипнет» еще одна клетка без глазка, то в ней установится концентрация 2,5, и так далее, каждый раз вдвое меньше. Раньше это свойство клеточных мембран Protozoon не имело существенного значения, но теперь оно становится ключевым фактором формообразования.
Обратите внимание, что в данном случае в созревшем двуклеточном организме (когда и передняя, и задняя клетка уже готовы поделиться) первой всегда делится передняя клетка. Это принципиальный момент. Из-за этого в задней клетке, которая теперь стала третьей по счету, концентрация X быстро снизилась до 2,5 — а при такой концентрации клетка делится уже не митозом, а мейозом.
Мы помним, что у исходного одноклеточного существа вещество X регулировало деление клеток: при концентрации выше четырех клетка делилась митозом, при меньшей концентрации — мейозом. Но для одноклеточного существа были безразличны небольшие индивидуальные различия в скорости начала клеточного деления. Между тем концентрация X (Px) немного влияла на эти скорости. В этом нет ничего удивительного, ведь в конечном счете в клетке все процессы хоть немного, но влияют друг на друга. По этому признаку — характеру влияния X на время начала деления — существовала индивидуальная изменчивость, но пока организм был одноклеточным, эти различия ни на что не влияли, и их было трудно даже заметить.
У той особи, из которой в результате мутации сформировался двуклеточный организм, митоз шел быстрее всего при концентрации X, близкой к 10. Поэтому первой стала делиться передняя клетка, а не задняя.
А что будет, если мутация «нерасхождения» произойдет у другой особи, у которой оптимум митоза случайно оказался ближе к Px = 5? Тогда строение многоклеточного организма окажется еще более сложным.

Взрослый организм теперь оказывается четырехклеточным! Он ведет себя как целое и обладает сложным детерминированным (предопределенным) онтогенезом, в ходе которого отдельные части эмбриона определенным образом влияют друг на друга — точно так же, как у настоящих эмбрионов. Такие взаимные воздействия называют «морфогенетическими корреляциями». Более того, онтогенез этого существа обладает еще одним характерным свойством «настоящего» онтогенеза — эквифинальностью, или помехоустойчивостью, то есть способностью компенсировать отдельные нарушения и все равно приходить к той же самой конечной цели.
Например, что будет, если приплывет хищная амеба и откусит у молодого четырехклеточного организма головную или хвостовую клетку? Нетрудно рассчитать, что уже через несколько шагов последствия травмы будут полностью компенсированы и жизненный цикл пойдет по исходному сценарию. То же самое справедливо и для стадии двухклеточного организма. Между прочим, недавние исследования показали, что у реальных животных регенерация — восстановление утраченных частей тела — управляется той же молекулярной «программой», которая руководит развитием этих частей в онтогенезе (см. сюжет «Разгадан механизм регенерации конечностей» в главе «Животные»). Кажется, наше выдуманное существо очень понятно объяснило нам, почему так получается и как это работает.
Но это еще далеко не все. В созревшем четырехклеточном организме мейоз генеративных клеток произошел раньше, чем митоз второй соматической клетки (у которой Px = 5). Но у других особей Metazoon (так мы назовем эти новые многоклеточные организмы) в такой ситуации митоз второй соматической клетки может произойти раньше, чем мейоз генеративных клеток. Как и в предыдущем случае, мы предполагаем, что все это было заложено в генофонде исходной популяции Protozoon в виде случайных вариаций, абсолютно несущественных для него (нейтральных), пока он был одноклеточным. Кто же мог знать, что такие пустяки вдруг станут жизненно важными в будущем.
—————Не забывайте об изменчивости! Как недавно выяснили биологи — преподаватели вузов, одна из основных причин недопонимания эволюционных идей у студентов состоит в том, что многие из них на интуитивном уровне считают всех представителей одного и того же вида одинаковыми. А преподаватели порой забывают в явном виде указать им, что это не так. Я тоже чуть не забыл. Так вот, очень важно всегда помнить, что каждая особь уникальна, все живые существа хоть немножко, но разные. Поэтому отбору всегда есть из чего выбирать. Изменчивость живых организмов, относящихся к одному и тому же виду, связана с тем, что многие гены в популяции существуют в виде нескольких вариантов (их называют аллельными вариантами, или аллелями). Набор генов у каждого представителя данного вида один и тот же, а вот комбинации аллелей у всех разные.—————
Итак, рассмотрим Metazoon, у которого при Px = 5 митоз происходит особенно быстро (так, что не успевает произойти мейоз в одновременно созревших клетках). В этом случае неизбежно возникает восьмиклеточный организм, а заодно и новый способ бесполого размножения (на рисунке все идет по часовой стрелке).
Фокус тут в том, что, когда клеток в цепочке становится пять, в последней из них Px снижается настолько, что образуется глазок! И вся картина сразу меняется.
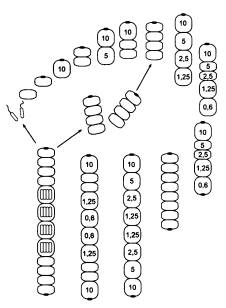
Как видим, двухклеточная фаза выпала из основного цикла. Теперь она осталась только в «раннем онтогенезе». После выхода гамет образуются два молодых четырехклеточных организма (бесполое размножение).
Но и это еще не все — нашу модель можно достраивать и дальше, производя (мысленно) все более и более сложные многоклеточные организмы на основе простых и вполне реалистичных допущений о характере нейтральной изменчивости у исходного Protozoon.
В последнем рассмотренном случае, когда в двух центральных клетках организма (на стадии 10 клеток) Px упала ниже критического уровня 1, глазок не образовался. Видимо, это произошло потому, что механизм образования глазка не срабатывает, если клетка контактирует более чем с одной соседней клеткой.
Однако вполне можно допустить, что среди исходных Protozoon, опять же в порядке нейтральной и безразличной изменчивости, были и такие особи, у которых механизм образования глазка все-таки мог бы сработать в этой ситуации. Из такого Protozoon, если у него произойдет мутация «нерасхождения», получатся уже не нитчатые (одномерные) а плоские (двумерные) многоклеточные организмы.
Здесь глазок появляется только в одной из двух центральных клеток, где Px упала до 0,6, поскольку во второй после этого Px сразу резко возрастает.
Крестообразный десятиклеточный организм становится отныне центральной фигурой, а все предшествующие этапы и циклы — его «ранним онтогенезом». Не перерисовывая эти стадии, проследим дальнейшую, строго детерминированную судьбу нашего десятиклеточного двуглазого Metazoon.
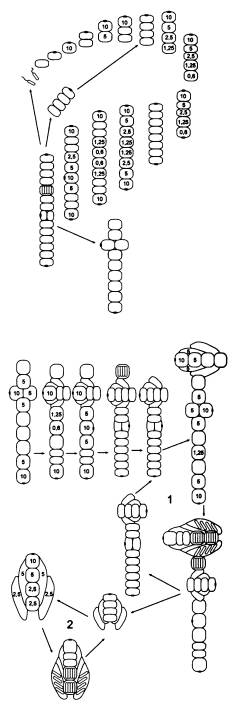
Как видим, получился очень интересный онтогенез с двумя замкнутыми циклами. В первом цикле основная жизненная форма — сложный восемнадцатиклеточный организм, который, созрев, производит гаметы, воспроизводит точную уменьшенную копию самого себя и одновременно отпочковывает восьмиклеточный двусторонне-симметричный организм с характерным обликом. Второй цикл состоит в том, что этот восьмиклеточный организм, произведя гаметы, автоматически возвращается к своему исходному строению.
Вот такое разнообразие сложных организмов, циклов и онтогенезов образуется само собой из вполне обычного простейшего Protozoon в результате одного небольшого (и очень вероятного) изменения — клетки перестали расходиться после митоза. Ясно, что здесь просчитаны и нарисованы далеко не все возможные варианты. Если допустить, что исходная изменчивость Protozoon по таким не очень важным для него признакам, как пороговые концентрации Px, вероятность образования глазка в разных непредусмотренных одноклеточным организмом ситуациях, реакция на внешние условия и др., достаточно велика, то число вариантов многоклеточных форм становится очень большим. Однако каждый из вариантов строго детерминирован при данном наборе параметров.
Вполне возможно, что в этом разнообразии найдется что «подхватить» отбору. Лишние стадии (например, часть мейозов) могут впоследствии выпасть, заменившись, допустим, апоптозами (смертью клеток); регуляторные механизмы могут усовершенствоваться и даже вовсе замениться; «головные» вегетативные отделы могут постепенно трансформироваться в настоящие головы, взяв на себя особые функции (при этом глазок и индуцируемые им изменения активности генов могут лечь в основу более совершенной системы регуляции онтогенеза).
Хочется обратить внимание на интересную особенность восьмиклеточного организма, образующего цикл 2 (на последнем рисунке): пространственное расположение клеток и их плотное «прилипание» друг к другу привели к тому, что клетки дифференцировались на два четких типа, напоминающие настоящие ткани: округлые осевые клетки и сильно вытянутые боковые клетки. Эти последние по своему расположению и форме явно имеют предпосылки в будущем превратиться в покровную ткань (эпителий). Кроме того, в цикле 2 решающую роль играет передача вытянутыми боковыми клетками сигнального вещества X от головной клетки — задним осевым клеткам. Это стабилизирует систему индивидуального развития, выравнивая величину Px вдоль продольной оси тела и предотвращая образование глазка на заднем конце. Такая функция продольных клеток создает предпосылки для того, чтобы в будущем на этой основе начала развиваться нервная система.
В нашей модели (как и в реальном развитии многоклеточных органзмов) каждая отдельная клетка имеет одну и ту же заложенную в нее изначально программу, содержащую несколько условных переходов («если — то»). Но это вовсе не «программа развития организма», а всего лишь программа поведения отдельной клетки. Совместно развиваясь, клетки меняют входные параметры не только у себя самих, но и у других клеток развивающегося организма, заставляя их тоже менять свое поведение. Именно в этом взаимном влиянии клеток развивающийся организм проявляет себя как целое. Из индивидуальных межклеточных взаимодействий каким-то чудесным образом, без всякого изначального плана и разумного замысла, рождается принципиально новая информация — программа развития организма.
Мне эта мысль представляется очень важной для понимания законов развития. И у Metazoon, и в онтогенезе настоящих многоклеточных все происходит настолько закономерно и целенаправленно, что нам кажется, будто развитие идет в соответствии с неким единым, заранее продуманным планом. Но мы видели, что ни у Protozoon, ни у разнообразных произошедших от него Metazoon никакой изначально заложенной программы развития нет и в помине. Эта «программа» рождается сама собой в процессе деления клеток словно бы из ничего. Не так ли обстоит дело и в случае настоящего онтогенеза? А в эволюции?
Главный вывод, который позволяет нам сделать гипотетический Protozoon и его чудесные превращения, можно сформулировать так: простая живая система может содержать в себе скрытые возможности самопроизвольного усложнения.
Опыты, подобные нашему мысленному эксперименту с Protozoon, сейчас активно проводятся и на реальных живых существах. Замечательным объектом такого рода оказалась почвенная бактерия Pseudomonas fluorescens.
—————Появлению многоклеточности мешают обманщики. Экспериментальное изучение эволюции бактерий — одно из перспективных направлений современной микробиологии. Пол Рейни из Оклендского университета (Новая Зеландия) является крупным специалистом в этой области. Один из его любимых объектов — бактерия Pseudomonas fluorescens из группы гамма-протеобактерий. Этот микроб, если предоставить ему необходимый минимум условий, охотно эволюционирует прямо на глазах у исследователей, осваивая новые ниши и вырабатывая различные оригинальные адаптации.В жидкой питательной среде бактерии развиваются сначала как одиночные, подвижные клетки, постепенно занимая всю толщу бульона. Когда в среде становится мало кислорода, получают преимущество бактерии-мутанты, образующие пленку на поверхности среды. Наблюдения за развитием таких колоний навели Рейни на любопытные идеи по поводу происхождения многоклеточности.В многоклеточном организме большинство клеток (так называемые соматические клетки) не передают своих генов следующим поколениям. Они размножаются делением, но только как часть целого организма, который рано или поздно погибает. Соматические клетки — своеобразный «эволюционный тупик», если смотреть на ситуацию с точки зрения индивидуальных клеток. Они жертвуют личными интересами для блага целого — точно так же, как рабочие особи у общественных насекомых.
Pseudomonas fluorescens — подвижная почвенная бактерия, помогающая биологам изучать эволюцию «в пробирке».
Вопрос в том, каким образом популяция индивидуальных клеток могла превратиться в единую систему, размножающуюся как целое. Почему естественный отбор перестал эффективно работать на уровне индивидуальных клеток и стал действовать на уровне клеточных популяций?В основе этого превращения, конечно, лежит кооперация между клетками, основанная на том, что в определенных ситуациях индивидууму становится выгодно немного поступиться сиюминутными личными интересами ради коллектива. У Pseudomonas «коллектив» образуется из бактерий-мутантов, которые выделяют повышенное количество веществ, способствующих склеиванию клеток. Такие бактерии после деления не могут «отклеиться» друг от друга. Фокус тут в том, что одиночные клетки барахтаются в толще бульона, а склеившиеся всплывают на поверхность, где кислорода гораздо больше. В результате образуется пленка из бактерий на поверхности среды. Производство клея — дело дорогостоящее, однако общая награда (кислород) с лихвой покрывает расходы.
Колония бактерий-мутантов Pseudomonas fluorescens на поверхности питательной среды.
Возникновение подобных колоний — уже большое эволюционное достижение, но до настоящего многоклеточного организма тут еще далеко. Колонии недолговечны, а главное, неспособны размножаться как единое целое. Отбор в них по-прежнему действует на индивидуальном, а не на групповом уровне. Он благоприятствует клеткам-«жуликам», которые перестают производить клей, однако продолжают пользоваться преимуществами жизни в группе. Нет никаких механизмов, которые препятствовали бы такому жульничеству. Безнаказанность способствует быстрому размножению обманщиков, что вскоре приводит к разрушению колонии.Рейни предположил, что ключевым моментом в возникновении многоклеточного организма должна быть выработка механизма борьбы с клетками-обманщиками. Но как может возникнуть такой механизм, если не работает отбор на уровне групп? Чтобы заработал групповой отбор, колония должна научиться размножаться как единое целое — иными словами, должно произойти разделение клеток колонии на вегетативные (сому) и генеративные (герму). Колонии бактерий, о которых идет речь, представляют собой как бы одну сплошную сому, то есть эволюционный тупик.Таким образом, для возникновения многоклеточности должны быть выполнены три условия: 1. кооперация; 2. механизм борьбы с обманщиками; 3. коллективное размножение. Причем все три эволюционные инновации должны развиться более или менее одновременно, что кажется очень маловероятным. Рейни предлагает следующий сценарий выхода из тупика. Если колония, не способная размножаться как целое, подобна соме, то в клетках-обманщиках можно увидеть прообраз гермы. Хотя их размножение губительно для колонии, они в принципе могут взять на себя роль тех «семян», из которых будут развиваться новые колонии. Обманщики действительно могут уплыть из колонии, когда захотят, — ведь они не приклеены к ней в отличие от «честных» бактерий.Если бы обманщики могли давать начало новым колониям, это создало бы предпосылки для группового отбора, который, в свою очередь, смог бы контролировать количество производимых колонией обманщиков. Допустим, в одних колониях мутации, приводящие к возникновению обманщиков, происходят часто, а в других — редко. Если число обманщиков растет слишком быстро, колония будет очень недолговечной, а с ее распадом теряют все свои преимущества и сами обманщики. Поэтому такая колония в конечном счете произведет меньше обманщиков, то есть оставит меньше потомства, чем та, в которой скорость производства обманщиков ниже. Таким образом, отбор, действующий теперь на уровне групп, в принципе может контролировать темп производства обманщиков, и в результате они из эгоистичных паразитов превратятся в неотъемлемую часть целостного организма — генеративные клетки (герму).Чтобы предложенная модель работала, обманщики должны с высокой частотой мутировать обратно в клетки, выделяющие клей. Иначе никаких новых колоний из них не получится. В лабораторных популяциях, с которыми работает Рейни, такая способность у обманщиков имеется. Впрочем, совершенно ясно, что прогрессивное развитие многоклеточных форм не может далеко продвинуться на основе случайной мутационной изменчивости — пусть даже скорость и направленность появления мутаций регулируются отбором (направленность мутирования может состоять в том, что меняются или перестраиваются совершенно определенные гены, а не все подряд, — это явление широко распространено, — однако характер изменений в этих генах все равно остается случайным). Чтобы предложенная схема заработала эффективно, мутационная изменчивость должна смениться модификационной. Это значит, что клетки должны обрести способность становиться «честными» (соматическими) или «обманщиками» (генеративными) за счет регуляции активности генов, не внося наследуемых изменений в геном.Как справедливо отмечает Рейни, такое преобразование вполне по силам эволюции. Однако нужно добавить, что способности бактериальной клетки к модификационной изменчивости крайне ограниченны по сравнению с эукариотической (ядерной) клеткой. Это объясняется прежде всего тем, что у бактерий нет клеточного ядра и наследственный материал постоянно находится под биохимическим ударом цитоплазматических реакций (см. раздел «Сообщество превращается в организм» в главе «Великий симбиоз»). Так что выработать сложные и эффективные системы регуляции активности генов при помощи разнообразных специализированных регуляторных молекул бактериям очень трудно.Возможно, именно по этой причине прокариотам так и не удалось приобрести настоящую многоклеточность, хотя они многократно подходили буквально вплотную к этому. Эукариоты, по имеющимся оценкам, достигали многоклеточного уровня организации как минимум 24 раза независимо в разных эволюционных ветвях.(Источник: Paul B. Rainey. Unity from conflict // Nature. 2007. V. 446. P. 616.)—————



