Книга: Рождение сложности. Эволюционная биология сегодня: неожиданные открытия и новые вопросы
Назад: Сообщество превращается в организм
Дальше: Этапы великой драмы
Из кусочков
В настоящее время в научной литературе обсуждается два или три десятка возможных сценариев превращения сообщества прокариот в эукариотическую клетку Они разнятся в деталях, но имеют много общего в целом. Общепризнанными считаются следующие факты.
1. Митохондрии (органеллы эукариотической клетки, ответственные за кислородное дыхание) являются прямыми потомками прокариот из группы альфапротеобактерий. К этой группе бактерий относятся современные внутриклеточные паразиты риккетсии (такие как возбудитель сыпного тифа, а также вольбахия, о которой рассказано в главе «Наследуются ли приобретенные признаки?»), некоторые аноксигенные фототрофы — пурпурные бактерии — и многие другие микробы. В качестве неоспоримых доказательств своего бактериального происхождения митохондрии по сей день сохранили маленькую кольцевую хромосому прокариотического типа с несколькими функционирующими генами (все остальные митохондриальные гены перебазировались в ядро и стали частью «общего» ядерного генома клетки), способность к самостоятельному размножению путем деления (причем осуществляется это деление при помощи тех же самых белков, что и у свободноживущих бактерий), типично бактериальный аппарат синтеза белка и множество других признаков. Сомневаться не приходится: предки митохондрий, свободноживущие альфапротеобактерии, когда-то были «проглочены» другим одноклеточным организмом, но не переварились, а стали неотъемлемой частью клетки.
2. Пластиды (органеллы растительной клетки, отвечающие за фотосинтез) являются потомками цианобактерий. Сходство пластид со свободноживущими цианобактериями настолько велико, что никаких сомнений тут просто не может быть.
3. Что касается «всего остального», то есть цитоплазмы эукариотической клетки и ее ядра, то здесь наблюдается причудливое смешение признаков архей и некоторых групп бактерий, а также уникальных черт, свойственных только эукариотам.
Основные различия между имеющимися гипотезами происхождения эукариот касаются происхождения «ядерно-цитоплазматического компонента» эукариотической клетки. Ясно, что его предком был какой-то крупный одноклеточный организм, который «проглотил» сначала будущих митохондрий, а потом будущих пластид, и превратил их в своих внутренних «сожителей»-симбионтов. Или, может быть, они не были проглочены, а проникли в него по собственной инициативе (подобно тому, как сейчас это делают внутриклеточные паразитические бактерии). Проблема в том, что этот организм-хозяин, насколько мы можем судить, был не очень похож на современных, доживших до наших дней прокариот. Он обладал рядом уникальных свойств.
Существует несколько версий его происхождения. Одни эксперты считают, что это была архея, возможно, близкая к современным термоплазмам или ферроплазмам, о которых мы немного рассказали в главе «Происхождение жизни». Все уникальные свойства развились уже после приобретения внутренних симбионтов (митохондрий) и объединения разнородных геномов в единый ядерный геном. Согласно другой гипотезе, предком цитоплазмы и ядра эукариот был представитель не архей и не бактерий, а некоей особой вымершей группы прокариот. Согласно третьей точке зрения, это был химерный организм, образовавшийся в результате слияния клеток нескольких разных архей и бактерий. Впрочем, дело могло обойтись и без слияния — оказалось достаточно очень интенсивного обмена генами между разными микробами, чтобы их свойства перемешались в одной клетке (о горизонтальном генетическом обмене подробно рассказано в главе «Наследуются ли приобретенные признаки?»).
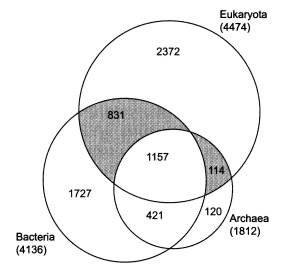
Распределение белковых семейств в трех надцарствах живой природы. Площади кругов соответствуют количеству семейств белков, встреченных у представителей данного надцарства. Как видно из рисунка, 1157 белковых семейств являются общими для всех трех надцарств, 2372 встречены только у эукариот, 831 есть у эукариот и бактерий, но не у архей, и т. д. (из статьи: Марков А. В., Куликов А. М. Происхождение эвкариот: выводы из анализа белковых гомологий в трех надцарствах живой природы // Палеонтол. журн. 2005. № 4. С. 3–18. ).
У каждой из версий, понятное дело, есть свои аргументы и свои сторонники. Лично мне больше всего нравится «химерная» теория. По крайней мере, именно к ней склонил нас с А. М. Куликовым (Институт биологии развития РАН) сравнительный анализ семейств белков, имеющихся у представителей трех надцарств живой природы — архей, бактерий и эукариот. Белки архейного происхождения, хотя их не очень много (114 семейств, см. рисунок), играют в эукариотической клетке ключевую роль. Именно они отвечают за работу с генетической информацией — транскрипцию, трансляцию, репликацию. Сюда относятся и НК-полимеразы, о которых шла речь выше, а также большинство белков, входящих в состав рибосом. Это позволяет предполагать, что в основе ядерно-цитоплазматического компонента эукариот лежала именно архея, а не бактерия и не какая-то особая «третья» группа прокариот, не дожившая до наших дней.
Многие белки бактериального происхождения попали в эукариотическую клетку вместе с «проглоченными» симбионтами — предками митохондрий и пластид. Однако среди «бактериальных» белков цитоплазмы и ядра есть и много таких, которые, скорее всего, не могли быть получены таким способом. Речь идет о тех семействах белков, которые есть у эукариот и есть также у бактерий, но не у тех, от которых произошли органеллы, а у каких-нибудь других. Иными словами, в эукариотической клетке существуют «бактериальные» семейства белков, которые не могли быть получены первыми эукариотами ни от «проглоченных» альфапротеобактерий, ни от цианобактерий — предков пластид. Но они, однако, могли быть получены от других бактерий — в особенности от различных бродильщиков (гетеротрофных бактерий, сбраживающих углеводы в бескислородных условиях). Похоже, именно от бродилыциков эукариоты получили, в частности, ферменты гликолиза — так называется важнейший энергетический процесс, происходящий в цитоплазме эукариотической клетки. Суть его в том, что молекула глюкозы расщепляется («сбраживается») без использования кислорода до пировиноградной кислоты (пирувата), и при этом происходит синтез АТФ. Пируват является для цитоплазмы конечным продуктом обмена, «отходом жизнедеятельности». Но пируват затем попадает в митохондрии, которые «сжигают» его в своей кислородной печке с огромным выходом энергии (которая тоже используется для синтеза АТФ). В совокупности бескислородный гликолиз, происходящий в цитоплазме, и кислородное дыхание, происходящее в митохондриях, являются главными источниками АТФ для эукариотической клетки.
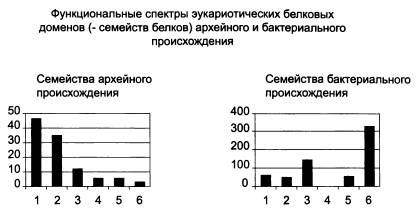
Распределение эукариотических белков архейного и бактериального происхождения по шести функциональным группам: 1 — синтез белка, 2 — репликация, транскрипция, модификация ДНК и РНК, 3 — сигнальные и регуляторные белки, 4 — образование мембранных пузырьков, 5 — транспортные и сортировочные белки, 6 — обмен веществ.
И вот на основе всех этих данных, а также с учетом того, что известно о строении и функционировании микробных сообществ, вырисовывается следующий возможный сценарий превращения сообщества прокариот в эукариотическую клетку (конечно, надо помнить, что это только один из возможных сценариев).
Назад: Сообщество превращается в организм
Дальше: Этапы великой драмы

