Книга: Рождение сложности. Эволюционная биология сегодня: неожиданные открытия и новые вопросы
Назад: Непрошеные помощники[21]
Дальше: Реликтовые микробные сообщества
От самодостаточных эгоистов к единой биосфере
Принято считать, что изначально в атмосфере Земли кислорода практически не было. Следовательно, железобактерии, с которыми мы познакомились в предыдущем разделе, не могли быть первыми, не с них началась на нашей планете «прокариотная эра».
Судя по результатам сравнительно-генетических исследований, да и по логике вещей, первыми или, по крайней мере, одними из первых могли быть другие хемоавтотрофы — археи-метаногены. В простейшем случае они получают энергию, восстанавливая углекислый газ до метана при помощи молекулярного водорода:
4H2+CO2 = CH4 + 2H2O.
Некоторое количество водорода образуется в земной коре в результате реакции перегретых горных пород с парами воды. Углекислый газ постоянно поступает из недр в атмосферу (например, при извержениях вулканов). Вот она, полная самодостаточность! Археям-метаногенам достаточно для жизни водорода, углекислого газа и воды (ну, конечно, в небольших количествах нужны еще азот, фосфор и всякие микроэлементы). Эти существа могли бы жить хоть на Марсе, хоть на полюсах Венеры, ведь они поразительно термостойки. Судя по сравнительно-генетическим данным, археи-метаногены появились около 3,8–4,1 млрд лет назад.
Незамкнутость цикла в данном случае создает угрозу накопления метана в атмосфере, что может привести к сильному парниковому эффекту. Однако активность метаногенов на древней Земле, возможно, была ограничена. Лимитирующим фактором мог служить, например, недостаток молекулярного водорода.
—————Самая главная химическая реакция. Жизнь на Земле основана на способности автотрофных организмов производить органику из углекислого газа (CO2). Поэтому тот химический процесс, в ходе которого CO2 включается в состав органических соединений, «фиксация CO2», по праву считается одним из важнейших (если не самым важным) в живой природе.Большинство организмов использует для фиксации CO2 циклическую последовательность химических реакций, известную под названием «цикл Кальвина». Ключевой фермент цикла Кальвина называется «рубиско» (это сокращенный вариант труднопроизносимого полного названия «рибулозобисфосфат карбоксилаза/оксигеназа»). Рубиско — самый распространенный в мире фермент. Он присоединяет молекулу CO2 к органическому веществу, которое называется рибулозобисфосфат (сокращенно — RuBP). Таким образом молекула углекислого газа оказывается включенной в состав органических соединений.Недавно выяснилось, что у архей способ фиксации CO2 отличается от классического цикла Кальвина. Главное отличие состоит в том, что у архей в этом процессе непосредственное участие принимают рибонуклеотиды. Это хорошо согласуется с теорией РНК-мира. Фиксация CO2 — один из древнейших биохимических процессов, который должен был появиться еще в то время, когда все функции в живых организмах выполнялись молекулами РНК, рибонуклеотидами и их производными. Поэтому следовало ожидать, что и фиксация CO2 изначально должна была идти под контролем и при активном участии этих молекул.В цикле Кальвина участвует специальный фермент, ответственный за синтез RuBP. Этот фермент есть у растений и автотрофных бактерий, однако у архей он отсутствует. Поэтому до самого последнего времени было неясно, откуда архей берут RuBP, вещество, абсолютно необходимое для фиксации CO2.Эту загадку удалось разгадать японским биохимикам в 2007 году. Оказалось, что RuBP образуется у архей не из других фосфорилированных сахаров, как в цикле Кальвина, а из молекулы, являющейся составным блоком РНК и выполняющей «по совместительству» множество других функций в живой клетке, — аденозинмонофосфата (АМФ). АМФ — это рибонуклеотид, состоящий, как и все рибонуклеотиды, из азотистого основания (в данном случае аденина), сахара (рибозы) и одного остатка фосфорной кислоты.Есть все основания полагать, что этот способ фиксации CO2 эволюционно древнее, чем цикл Кальвина. Не случайно он обнаружен именно у архей — организмов, которых многие специалисты считают самыми архаичными формами жизни.(Источник: Takaaki Sato, Haruyuki Atomi, Tadayuki Imanaka. Archaeal Type III RuBisCOs Function in a Pathway for AMP Metabolism // Science. 2007. V. 315. P. 1003–1006.)—————
В течение первого миллиарда лет существования прокариотной биосферы (примерно от 4,1 до 3,2 млрд лет назад) одни за другими появлялись новые формы микробов и новые способы получения энергии. В числе первых, по-видимому, появился бескислородный (аноксигенный) фотосинтез (? — 3,2–3,7 млрд лет назад). Его освоили бактерии — предки нынешних зеленых, пурпурных и цианобактерий.
Аноксигенные фотосинтезирующие бактерии научились использовать энергию солнечного света при помощи особых светочувствительных молекул — бактериохлорофиллов и связанных с ними белковых комплексов. Молекулярная «машина», необходимая для осуществления эффективного аноксигенного фотосинтеза, устроена довольно сложно (хотя и намного проще, чем та, что используется оксигенными фотосинтетиками, появившимися позже). По-видимому, эффективный фотосинтез возник не сразу. Начиналось все с того, что некоторые микробы научились немного «подкармливаться» солнечным светом при недостатке других источников энергии. Для такой «подкормки» не нужны сложные молекулярные комплексы — достаточно одного-единственного светочувствительного белка. Недавно выяснилось, что подобные архаичные, простые и малоэффективные способы утилизации солнечного света до сих пор очень широко распространены в мире прокариот. Стало ясно, что способность к росту за счет энергии солнечного света (фототрофность) распространена в живой природе гораздо шире, чем считалось до сих пор.
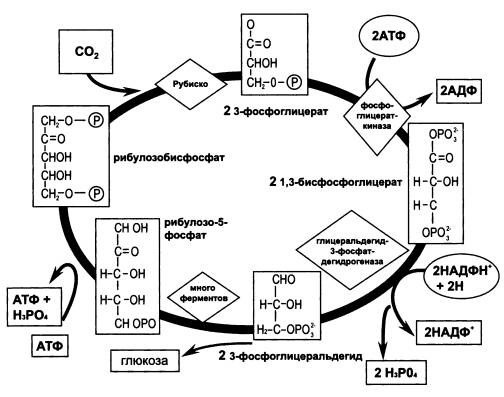
Цикл Кальвина — важнейший биохимический процесс в живой природе, в ходе которого неорганический углерод вводится в состав органических молекул.
—————При недостатке растворенной органики морские бактерии подкармливаются солнечным светом. Открытия последних лет показали, что кроме зеленых растений и цианобактерий, осуществляющих фотосинтез при помощи хлорофилла а, и известных ранее фототрофных бактерий, делающих то же самое при помощи бактериохлорофиллов, питаться солнечным светом могут и многие другие микробы, обладающие светочувствительными белками — протеородопсинами.Протеородопсины представляют собой своеобразные «насосы», локализованные в клеточной мембране и использующие энергию солнечного света для перекачки протонов (H+) из клетки во внешнюю среду. Таким образом световая энергия переводится в разность электрохимических потенциалов, которая может затем использоваться клеткой для различных нужд, в том числе для синтеза АТФ. АТФ в свою очередь является универсальной энергетической «разменной монетой» клетки, используемой в самых разных энергоемких процессах.Протеородопсины были обнаружены у морских бактерий в 2000 году. Открытие привлекло большое внимание, поскольку оно означало, что многие организмы, до сих пор считавшиеся строго хемотрофными (получающими энергию за счет химических реакций), в действительности могут оказаться фототрофными — по крайней мере отчасти. В таком случае все представления об энергетическом балансе биосферы нуждаются в пересмотре.Фототрофность на основе протеородопсинов — это гораздо менее эффективный способ утилизации солнечной энергии по сравнению с настоящим фотосинтезом, но зато и гораздо более простой.Вскоре выяснилось, что протеородопсины встречаются не только у бактерий, но и у многих архей, обитающих в морях и океанах. По-видимому, морские микроорганизмы активно обмениваются друг с другом генами протеородопсинов, которые служат им важным подспорьем в конкурентной борьбе за энергетические ресурсы. Простота протеородопсиновой системы утилизации солнечного света облегчает ее горизонтальную передачу от одних микробов другим (о горизонтальном обмене генами подробно рассказано в главе «Наследуются ли приобретенные признаки?»).Однако до сих пор никому не удавалось экспериментально показать, что морские микробы, геном которых содержит гены протеородопсинов, действительно являются фототрофными организмами, то есть могут использовать энергию солнечного света для роста. Более того, первые попытки это подтвердить дали отрицательный результат: массовая и вездесущая морская бактерия Pelagibacter ubique, имеющая протеородопсины в клеточной мембране, в лабораторных условиях росла на свету не лучше, чем в темноте. Проблема осложнялась еще и тем, что большинство морских бактерий, содержащих протеородопсины, относятся к числу некультивируемых — в лабораторных условиях они просто не живут. Поэтому оставалось неопровергнутым предположение, что протеородопсиновая система использования солнечного света, возможно, настолько неэффективна, что не может внести заметного вклада в энергетический баланс морских экосистем и используется микробами лишь для каких-то второстепенных нужд.Лишь в 2007 году микробиологам из Швеции и Испании удалось показать, что это не так. Они обнаружили протеородопсины у бактерии Dokdonia, обитающей в Средиземном море. Бактерию удалось культивировать в лабораторных условиях. Оказалось, что в натуральной морской воде бактерия на свету растет гораздо лучше, чем в темноте. В первом случае плотность популяции после 100 часов инкубации составляла 3 x 105 клеток на миллилитр, во втором — в шесть раз меньше (0,5 x 105). Если культуру, выращенную в темноте, осветить, бактерии начинают быстро размножаться; если оставить в темноте — их численность постепенно снижается. Кроме того, бактерии, выросшие на свету, заметно крупнее тех, что росли в потемках.Дополнительные эксперименты показали, что влияние света на рост бактерий зависит от концентрации растворенной органики. Конечно, протеородопсины в отличие от систем настоящего фотосинтеза не могут сделать бактерию полностью автотрофной, то есть не нуждающейся в готовых органических веществах. Такие бактерии лишь «подкармливаются» светом и без готовой органики существовать не могут. Dokdonia практически не растет в воде с содержанием органики ниже определенного порога, и свет нисколько не помогает ей в этой ситуации. Однако при более высоких концентрациях растворенного органического вещества бактерия растет на свету в несколько раз быстрее, чем в темноте. Если же поместить докдонию в воду с избытком органики, то свет снова перестает влиять на ее рост (в этих условиях она растет одинаково хорошо как на свету, так и в темноте).Таким образом, протеородопсины повышают жизнеспособность морских бактерий лишь в условиях низких (но не слишком низких) концентраций растворенной органики. Возможно, отрицательные результаты, полученные ранее с Pelagibacter, объясняются именно нерациональным количеством органики в опытных средах.Так или иначе, мы теперь знаем, что многочисленные и разнообразные морские микробы, имеющие в своем геноме гены протеородопсинов, действительно могут быть факультативными фототрофами, то есть способны разнообразить свою диету, состоящую в основном из растворенной органики, солнечным светом.(Источники: 1. Laura Gómez-Consarnau et al. Light stimulates growth of proteorhodopsincontaining marine Flavobacteria // Nature. 2007. V. 445. P. 210–213; 2. Gazalah Sabehi et al. New insights into Metabolic Properties of Marine Bacteria Encoding Proteorhodopsins // PLoS Biol. 2005. 3(8): e273.)—————
Изобретение аноксигенного фотосинтеза было большим шагом вперед. Живые существа, овладевшие секретом фотосинтеза, получили доступ к неисчерпаемому источнику энергии — солнечному свету. Правда, их зависимость от дефицитных химических веществ, поступающих понемногу из земных недр, при этом все-таки сохранилась. Дело в том, что для фотосинтеза одного света мало — нужно еще какое-нибудь вещество, от которого можно оторвать электрон (это называется «фотоокисление»). В простейшем случае в роли донора электрона при фотосинтезе выступает сероводород. В результате деятельности аноксигенных фотосинтетиков сероводород превращается в серу (S) или сульфат (SO42-). Опять незамкнутый цикл и накопление отходов!
Но жизнь уже набирала силу, разнообразие микробов росло, и незамкнутые циклы постепенно начинали замыкаться. Планета захлебывается метаном и сульфатами? Что ж, эволюция нашла отличный выход из сложной ситуации: появились микроорганизмы, способные окислять метан при помощи сульфатов. Это были не просто микробы, а симбиотические микробные сообщества, состоящие из архей и бактерий. Архей окисляли метан, а бактерии восстанавливали сульфаты, причем оба процесса были каким-то не до конца еще понятным образом сопряжены между собой в неразрывное целое. Такие сообщества сохранились и по сей день в соответствующих местах обитания — там, где достаточно метана и сульфатов (например, в окрестностях подводных грязевых вулканов — см. ниже сюжет «В подводном грязевом вулкане обнаружены неизвестные микробы»).
Результатом окисления метана был углекислый газ, необходимый всем автотрофам, а результатом восстановления сульфатов — сероводород, который с удовольствием использовали фотосинтетики. Циклы замыкались, биосфера приобретала устойчивость и способность к саморегуляции. Начиналась эпоха Великого Содружества Микробов.
Примерно в тот же период (свыше 3,2 млрд лет назад), по- видимому, появились и первые гетеротрофы — так называемые бродильщики, которые получают энергию за счет бескислородной ферментации (сбраживания) готовой органики, произведенной автотрофами. В качестве отходов жизнедеятельности бродильщики имеют обыкновение выделять молекулярный водород, до которого в древней биосфере уже были охотники: во-первых, археи-метаногены, во-вторых, бактерии-сульфатредукторы (они охотно используют молекулярный водород в качестве восстановителя для восстановления сульфатов).
Ясно, что на этом этапе большинство микроорганизмов уже не могли обходиться друг без друга. Даже в наши дни многие бродильщики наотрез отказываются расти в отсутствие микробов, утилизирующих выделяемый ими водород (сульфатредукторов или метаногенов), а тем, в свою очередь, жизнь не мила без бродилыциков.
Уже 3,55 млрд лет назад на Земле, по-видимому, существовали сложные микробные сообщества — бактериальные маты. Именно они, скорее всего, ответственны за образование древнейших строматолитов. В наши дни нечто подобное можно наблюдать в некоторых экстремальных местообитаниях, таких как горячие источники. Древние бактериальные маты, вероятно, состояли из двух слоев. В верхнем обитали аноксигенные фототрофы. Они синтезировали органику из углекислого газа, потребляли сероводород и выделяли сульфаты. В нижнем слое жили бродильщики (они потребляли органику, произведенную фототрофами, и выделяли водород), сульфатредукторы (потребляли сульфаты и водород, производили сероводород), а также, возможно, метаногены с метанотрофами. В ходе жизнедеятельности сообщества под ним постепенно, слой за слоем, накапливался уплотненный осадок — так формировались слоистые образования, известные под названием строматолитов. Карбонат кальция — основной строительный материал строматолита — отчасти осаждался из морской воды, отчасти продуцировался самими микробами (в первую очередь сульфатредукторами).
Назад: Непрошеные помощники[21]
Дальше: Реликтовые микробные сообщества

