Книга: В поисках памяти
Назад: 15. Биологические основы индивидуальности
Дальше: 17. Долговременная память
16. Молекулы и кратковременная память
В 1975 году, через двадцать лет после того, как Гарри Грундфест сказал мне, что мозг нужно исследовать по одной клетке, вместе с коллегами я приступил к изучению клеточных основ памяти, то есть механизмов, позволяющих нам на всю жизнь запоминать встречу с другим человеком, пейзаж, лекцию или диагноз. Нам удалось узнать, что память обеспечивается изменениями синапсов в нейронных цепях: кратковременная — функциональными изменениями, долговременная — структурными. Теперь нам хотелось проникнуть в тайну памяти еще глубже. Мы надеялись разобраться в молекулярной биологии психического явления и узнать, какие именно молекулы ответственны за кратковременную память. Задавшись этим вопросом, мы попали в совершенно неизведанную область.
Это предприятие казалось мне не таким уж безнадежным благодаря растущей уверенности в том, что аплизия — вполне подходящий простой объект для изучения молекулярных основ работы памяти. Мы проникли в лабиринт синаптических связей нервной системы аплизии, картировали нейронные проводящие пути ее рефлекса втягивания жабр и показали, что обеспечивающие этот рефлекс синапсы могут усиливаться в результате обучения. Мы в прямом смысле прокладывали путь по наружным кругам научного лабиринта. Теперь нам хотелось определить, где конкретно в исследованном нами проводящем пути происходят синаптические изменения, связанные с кратковременной памятью.
Мы сосредоточились на ключевой синаптической связи между сенсорным нейроном, передающим информацию о прикосновении к сифону моллюска, и мотонейроном, потенциал действия которого вызывает втягивание жабр. Нам хотелось узнать, какую роль играет каждый из двух нейронов, образующих эту связь, в связанном с обучением изменении синаптической силы. Изменяется ли в ответ на действие раздражителя сенсорный нейрон, так что окончание аксона выделяет больше или меньше нейромедиатора? Или изменение происходит в мотонейроне, так что на его мембране оказывается больше рецепторов нейромедиатора или повышается чувствительность этих рецепторов? Мы выяснили, что происходящие изменения носят вполне односторонний характер: при кратковременном привыкании, действие которого продолжается минуты, сенсорный нейрон выделяет меньше нейромедиатора, а при кратковременной сенсибилизации — больше.
Нейромедиатором, как мы впоследствии выяснили, в данном случае служит глутаминовая кислота (глутамат), которая также является важнейшим возбуждающим нейромедиатором головного мозга млекопитающих. Увеличивая количество глутамата, посылаемого сенсорным нейроном мотонейрону; сенсибилизация усиливает синаптический потенциал, возникающий при этом в мотонейроне, тем самым облегчая запускание потенциала действия, который приводит к втягиванию жабр.
Синаптический потенциал в моторном нейроне длится всего миллисекунды, но мы отметили, что удар током в заднюю часть тела аплизии усиливает выделение глутамата и тем самым синаптическую передачу на многие минуты. Как это происходит? Когда мы с коллегами сосредоточились на этом вопросе, мы заметили необычную вещь. Усиление синаптической связи между сенсорным нейроном и мотонейроном сопровождается очень медленным синаптическим потенциалом в сенсорной клетке, который длится минуты, а не миллисекунды, как это обычно бывает с синаптическим потенциалом в мотонейроне. Вскоре мы обнаружили, что удар током вызывает активацию другой разновидности сенсорных нейронов, получающих информацию от задней части тела аплизии. Эти сенсорные нейроны задней части тела активируют группу интернейронов, которые действуют на сенсорные нейроны, ведущие от сифона. Именно они и производят тот необычайно медленный синаптический потенциал. Затем мы задались следующими вопросами: какой нейромедиатор выделяют интернейроны? Как этот нейромедиатор вызывает выделение большего количества глутамата из окончаний сенсорного нейрона, тем самым обеспечивая хранение кратковременной памяти?
Мы обнаружили, что интернейроны, активируемые ударом тока в заднюю часть тела аплизии, выделяют нейромедиатор серотонин. Кроме того, эти интернейроны образуют синапсы и с телами клеток сенсорных нейронов, и с их пресинаптическими окончаниями и не только вызывают медленный синаптический потенциал, но и увеличивают выделение сенсорным нейроном глутамата в синаптическую щель, соединяющую его с мотонейроном. Более того, воздействуя серотонином на синаптические связи между сенсорными нейронами и мотонейронами, мы и сами могли вызывать медленный синаптический потенциал, увеличение синаптической силы и усиление рефлекса втягивания жабр.
Мы назвали эти выделяющие серотонин интернейроны модуляторными интернейронами, потому что они служат не для непосредственного обеспечения поведенческой реакции, а для модуляции рефлекса втягивания жабр за счет усиления связей сенсорных нейронов с мотонейронами.
Благодаря этим открытиям мы поняли, что существует два типа нейронных цепей, играющих важную роль в поведении и обучении: основные цепи, исследованные нами ранее, и модуляторные, которые мы еще только начинали подробно исследовать (рис. 16–1). Основные цепи служат для непосредственного обеспечения поведенческих реакций, значит, имеют кантианскую природу. Это предопределенные генетикой и механизмами развития нейронные компоненты поведения, его нейронное устройство. Основные цепи состоят из сенсорных нейронов, иннервирующих сифон, интернейронов и мотонейронов, управляющих рефлексом втягивания жабр. В ходе обучения основная цепь играет роль ученика, приобретающего новые знания. Модуляторные цепи, в свою очередь, имеют локковскую природу. Модуляторная цепь играет роль учителя. Она служит не для непосредственного обеспечения поведенческой реакции, но для точной настройки этой реакции в процессе обучения за счет гетеросинаптической модуляции силы связей сенсорных нейронов с мотонейронами. Модуляторная цепь, активируемая ударом тока в заднюю часть тела, учит аплизию обращать внимание на прикосновение к сифону, что может оказаться важным для ее безопасности. Таким образом, эта цепь, по сути, обеспечивает у аплизии состояние стресса, подобно аналогичным модуляторным цепям, которые служат неотъемлемыми компонентами механизма памяти у более сложных животных, в чем нам еще предстоит убедиться.
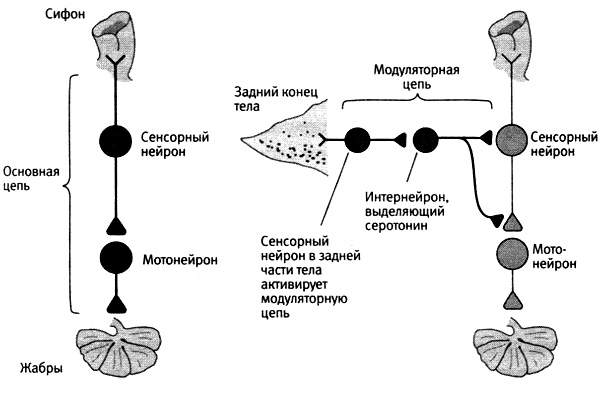
16–1. Два типа нейронных цепей в нервной системе. Основные цепи обеспечивают поведенческие реакции, а модуляторные воздействуют на основные, регулируя силу их синаптических связей.
Меня поразило, что роль модулятора сенсибилизации у аплизии играет именно серотонин! Одни из моих первых экспериментов, проведенных в 1956 году под руководством Дома Пурпуры, были посвящены действию серотонина. Более того, весной 1956 года в День студента в медицинской школе Нью-Йоркского университета я выступая с небольшим докладом на тему «Электрофизиология взаимодействия серотонина и ЛСД в афферентных проводящих путях коры головного мозга». Джимми Шварц любезно согласился выслушать репетицию этого доклада и помог мне его улучшить. Я начинал понимать цикличность жизни. Я не занимался серотонином почти двадцать лет и вот возвращался к нему с возобновленным интересом и увлеченностью.
Когда нам стало известно, что серотонин действует как модуляторный медиатор, увеличивая выделение глутамата из пресинаптических окончаний сенсорного нейрона, все было готово для биохимического исследования работы памяти. По счастью, на этом пути у меня был такой отличный проводник и напарник, как Джимми Шварц.
До своего возвращения в Нью-Йоркский университет Джимми работал в Рокфеллеровском университете с бактерией кишечной палочкой (Escherichia coli) — одноклеточным организмом, исследования которого позволили открыть многие фундаментальные принципы современной биохимии и молекулярной биологии. В 1966 году он переключился на аплизию и начал свои исследования этого организма с выявления химических медиаторов, выделяемых нейронами абдоминального ганглия. С 1971 года мы объединили усилия, чтобы исследовать молекулярные процессы, сопровождающие обучение.
Джимми оказал мне неоценимую помощь на этом втором большом этапе моего становления как биолога. На наши исследования повлияли работы Луиса Флекснера, показавшего за несколько лет до этого, что долговременная память у мышей и крыс требует синтеза новых белков, а кратковременная память не требует. Белки — главные работники клетки. Они составляют ее ферменты, ионные каналы, рецепторы и систему транспорта. Поскольку, как мы выяснили, в долговременной памяти задействовано образование новых связей, не было ничего удивительного в том, что для образования этих связей требуется синтез новых белков.
Мы с Джимми занялись проверкой этой идеи на аплизии — на уровне сенсорного нейрона сифона и его синапсов на мотонейронах жабр. Если синаптические изменения сопровождают изменения памяти, то выявленные нами кратковременные синаптические изменения не должны требовать синтеза новых белков. Именно это мы и обнаружили. Что же тогда обеспечивает эти кратковременные изменения?
Кахаль показал, что нервная система состоит из нейронов, специфическим образом связанных друг с другом проводящими путями. Я наблюдал эту удивительную специфичность связей в простых нейронных цепях, обеспечивающих рефлекторное поведение у аплизии. Но Джимми отметил, что эта специфичность распространяется и на молекулы — соединения атомов, которые служат функциональными элементами клетки. Биохимики установили, что молекулы могут взаимодействовать друг с другом в пределах клетки и что происходящие при этом химические реакции связаны в определенные последовательности, которые называют биохимическими сигнальными путями. Такие пути передают информацию в виде молекул от поверхности клетки в ее внутреннюю среду; подобно тому как нервные клетки передают информацию друг другу. Но это «беспроводные» пути. Молекулы, плавающие внутри клетки, распознаются другими специфическими молекулами и связываются с ними, регулируя их активность.
Мы не только реализовали мой давний замысел поймать выработанную в ходе обучения реакцию в наименьшей возможной популяции нейронов, мы также поймали один из компонентов простой формы памяти в единственной сенсорной клетке. Но даже в единственном нейроне аплизии содержатся тысячи различных белков и других молекул. Какие из них отвечают за кратковременную память? Когда мы с Джимми начали обсуждать, как это можно узнать, мы сосредоточились на идее, что серотонин, выделяемый в ответ на электрический удар, может увеличивать выделение глутамата из сенсорного нейрона, запуская в нем особую последовательность биохимических реакций.
Последовательность биохимических реакций, которую мы искали, должна была служить двум принципиальным целям. Во-первых, эти реакции должны были преобразовывать непродолжительное воздействие серотонина в молекулы, сигнал которых сохранялся бы внутри сенсорного нейрона в течение минут. Во-вторых, молекулы должны были передавать сигнал от клеточной мембраны, на которую действует серотонин, во внутреннюю среду клетки — в особые участки окончаний аксона, задействованные в выделении глутамата. Мы подробно изложили эти мысли в своей статье 1971 года, опубликованной в Journal of Neurophysiology, и высказали предположение, что в этом процессе могут быть задействованы молекулы одного особого вещества — так называемого циклического АМФ.
Что такое циклический АМФ? И почему мы сочли его вероятным претендентом на эту роль? Мне пришел в голову именно циклический АМФ в связи с тем, что было известно: небольшие молекулы этого вещества служат важнейшими регуляторами передачи сигналов в мышечных и жировых клетках. Мы с Джимми знали, что природа консервативна, поэтому механизм, используемый в клетках одной ткани, с большой вероятностью сохранится и будет использоваться в клетках другой. Эрл Сазерленд из Западного резервного университета Кейса в Кливленде в то время уже обнаружил, что гормон адреналин (эпинефрин) вызывает непродолжительные биохимические изменения на поверхности мембраны жировых и мышечных клеток, приводя к более продолжительным изменениям внутри клеток. Эти более продолжительные изменения происходят за счет повышения содержания циклического АМФ во внутренней среде клеток.
Революционные открытия Сазерленда были впоследствии описаны так называемой теорией вторичных посредников (вторичных мессенджеров). Основой этой теории биохимической передачи сигналов в клетках стало открытие на поверхности жировых и мышечных клеток нового класса рецепторов, реагирующих на гормоны. Бернард Кац ранее описал медиатор-зависимые рецепторы, которые относятся к так называемым ионотропным рецепторам. Когда с таким рецептором связывается нейромедиатор, он открывает или закрывает ворота проходящего сквозь этот рецептор ионного канала, тем самым преобразуя химический сигнал в электрический. Но в рецепторах нового типа, так называемых метаботропных, нет ионного канала, который они могли бы открывать и закрывать. Один участок этих рецепторов, выступающий из наружной поверхности клеточной мембраны, опознает сигналы, приходящие от других клеток, а второй участок, выступающий из внутренней поверхности мембраны, запускает работу определенного фермента. Когда такие рецепторы опознают и связывают молекулы химического посредника из внеклеточной жидкости, они активируют работающий внутри клетки фермент аденилатциклазу, который вырабатывает циклический АМФ.
У этого процесса есть преимущество — он позволяет многократно усиливать клеточную реакцию. Когда одна молекула химического посредника связывается с метаботропным рецептором, он активирует аденилатциклазу, которая производит тысячи молекул циклического АМФ. Затем циклический АМФ связывается с особыми белками, запускающими во всей клетке целый ряд молекулярных реакций. При этом аденилатциклаза продолжает вырабатывать циклический АМФ в течение минут. Поэтому метаботропные рецепторы обычно действуют сильнее, шире и дольше, чем ионотропные. Действие ионотропных рецепторов обычно продолжается миллисекунды, а действие метаботропных — секунды или минуты, то есть в тысячи или десятки тысяч раз дольше.
Чтобы отличать две разделенные в пространстве функции метаботропных рецепторов, Сазерленд назвал химический посредник, который связывается с рецептором на наружной стороне клеточной мембраны, первичным, а циклический АМФ, вырабатываемый внутри клетки для передачи сигнала, вторичным. Сазерленд доказывал, что вторичный посредник передает внутрь клетки сигнал, поступающий от первичного, на мембрану и вызывает реакцию на этот сигнал по всей клетке (рис. 16–2). Представление о вторичных посредниках заставило нас предположить, что метаботропные рецепторы и циклический АМФ могут оказаться теми неуловимыми факторами, которые обеспечивают связь медленного синаптического потенциала сенсорных нейронов с усиленным выделением глутамата, тем самым обеспечивая работу кратковременной памяти.
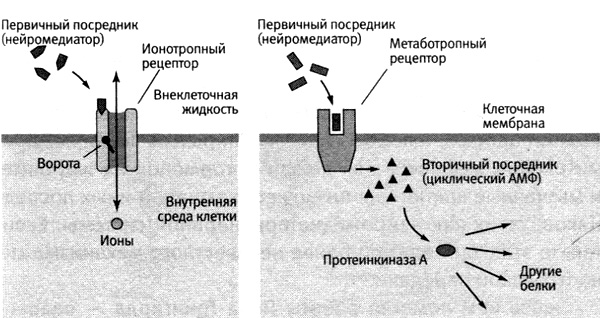
16–2. Два типа рецепторов, выделенных Сазерлендом. Ионотропные рецепторы (слева) вызывают изменения, которые длятся миллисекунды. Метаботропные рецепторы (например, рецепторы серотонина) действуют через вторичных посредников (справа). Они вызывают изменения, которые длятся секунды или минуты и передаются по всей клетке.
В 1968 году Эдвин Кребс из Вашингтонского университета получил первые данные о том, как циклический АМФ производит свое широкое действие. Циклический АМФ связывается с ферментом, который Кребс назвал АМФ-зависимой протеинкиназой или протеинкиназой А (потому что это была первая известная протеинкиназа). Киназы видоизменяют белки, присоединяя к ним молекулу фосфата; этот процесс называют фосфорилированием. Фосфорилирование активирует одни белки, но инактивирует другие. Кребс обнаружил, что фосфорилирование вполне обратимо и может служить простым молекулярным переключателем, включая и выключая биохимическую активность белка.
Затем Кребс стал разбираться с тем, как работает этот молекулярный переключатель. Он открыл, что протеинкиназа А представляет собой сложную молекулу, состоящую из четырех единиц — двух регуляторных и двух каталитических. Каталитические единицы устроены так, что могут осуществлять фосфорилирование, но регуляторные обычно «сидят» на них, подавляя их работу. У регуляторных единиц имеются участки, которые связывают циклический АМФ. Когда концентрация циклического АМФ в клетке увеличивается, регуляторные единицы протеинкиназы А связывают избыточные молекулы. Это приводит к изменению их формы, вызывая отпадение от каталитических единиц, которые получают возможность фосфорилировать белки-мишени.
Представление об этом помогло нам задаться ключевым вопросом, специфичен ли открытый Сазерлендом и Кребсом механизм для действия гормонов на жировые и мышечные клетки или он может включать и иных посредников, таких как нейромедиаторы нервной системы. Если так, то это будет пример ранее неизвестного механизма синаптической передачи.
Здесь нам помогли работы Пола Грингарда — одаренного биохимика, получившего также физиологическое образование, который недавно перешел в Йельский университет с должности директора отделения биохимии исследовательских лабораторий фармацевтической компании Geigy. По дороге в Йель он задержался на год на отделении Сазерленда. Осознав важность возможного нового механизма передачи сигналов в нервной системе, в 1970 году Грингард начал разбираться с метаботропными рецепторами в мозгу крысы. Тут произошло одно замечательное совпадение, связавшее Арвида Карлссона, Пола Грингарда и меня на нашем научном пути, который в 2000 году привел нас троих в Стокгольм, где мы разделили Нобелевскую премию по физиологии и медицине за исследование передачи сигналов в нервной системе.
В 1958 году Арвид Карлссон, великий шведский фармаколог, открыл, что дофамин играет в нервной системе роль медиатора. Затем он показал, что, когда в организме кролика снижается концентрация дофамина, у животного развиваются симптомы, напоминающие болезнь Паркинсона. Когда Грингард стал изучать метаботропные рецепторы головного мозга, он начал с рецептора дофамина и обнаружил, что этот рецептор включает фермент, который увеличивает содержание циклического АМФ и активирует протеинкиназу А в мозгу!
Пойдя по их стопам, мы с Джимми Шварцем открыли, что серотонин в ходе сенсибилизации тоже запускает работу циклического АМФ как вторичного посредника. Как мы уже убедились, удар током по задней части тела аплизии вызывает активацию модуляторных иитернейронов, которые выделяют серотонин. Серотонин, в свою очередь, на несколько минут увеличивает синтез циклического АМФ в пресинаптических окончаниях сенсорных нейронов (рис. 16–3). Итак, все сходилось: концентрация циклического АМФ остается повышенной примерно столько же, сколько длятся медленный синаптический потенциал, повышение синаптической силы связи сенсорных нейронов с мотонейронами и усиленная поведенческая реакция животного, вызванная электрическим ударом по его телу.
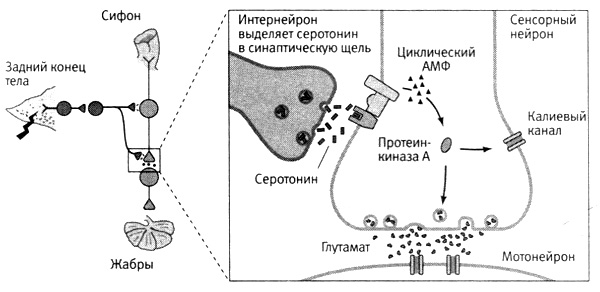
16–3. Этапы работы биохимического механизма долгосрочной памяти. Электрический удар по задней части тела аплизии вызывает активацию иитернейрона, который выделяет в синаптическую щель химический посредник серотонин. Преодолев синаптическую щель, серотонин связывается с рецептором на мембране сенсорного нейрона, запуская синтез циклического АМФ (1). Циклический АМФ вызывает отделение каталитической единицы протеинкиназы А (2). Каталитическая единица протеинкиназы А усиливает выделение нейромедиатора глутамата (3).
Первое прямое подтверждение того, что циклический АМФ задействован в формировании кратковременной памяти, было получено в 1976 году, после того как в нашу лабораторию пришел постдок из Италии Марчелло Брунелли. Брунелли осуществил проверку того, что повышение концентрации циклического АМФ в сенсорных нейронах, сигналом для которого служит серотонин, приводит к увеличению количества глутамата, выделяемого из окончаний этих нейронов. Мы вводили циклический АМФ непосредственно в сенсорный нейрон аплизии и обнаружили, что это вызывало резкое повышение количества выделяемого глутамата, а следовательно, и силу синаптической связи сенсорного нейрона с мотонейронами. Более того, введение в клетку циклического АМФ вызывало такое же повышение синаптической силы, как при воздействии серотонином на сенсорные нейроны или при воздействии током на заднюю часть тела животного. Этот замечательный эксперимент не только показал роль циклического АМФ в кратковременной памяти, но и дал нам первые сведения о молекулярных механизмах обучения. Теперь, когда мы научились улавливать основные молекулярные компоненты кратковременной памяти, мы могли использовать их для искусственной имитации формирования памяти.
В 1978 году мы с Джимми начали сотрудничать с Грингардом. Нам хотелось узнать, действует ли циклический АМФ на кратковременную память посредством протеинкиназы А. Мы разделили этот белок на составляющие и ввели непосредственно в сенсорный нейрон только каталитическую единицу, которая в норме и осуществляет фосфорилироеание. Выяснилось, что эта единица делает то же, что циклический АМФ: усиливает синаптическую связь, увеличивая выделение глутамата. Затем, чтобы лишний раз убедиться, что мы на верном пути, мы ввели в сенсорный нейрон ингибитор протеинкиназы А и обнаружили, что он действительно препятствует увеличению выделения глутамата под действием серотонина. Заметив, что присутствие циклического АМФ и протеинкиназы А не только необходимо, но и достаточно для усиления связи сенсорного нейрона с мотонейронами, мы смогли выявить первые звенья цепи биохимических событий, ведущих к формированию кратковременной памяти (рис. 16–4).
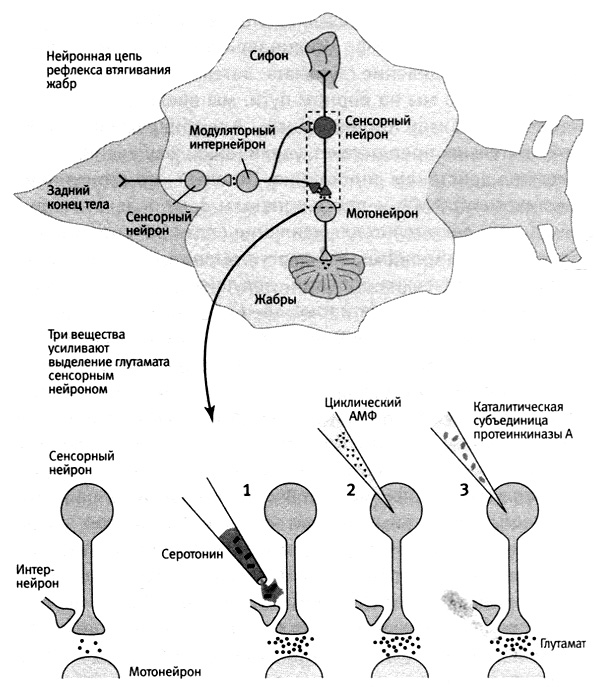
16–4. Молекулы, задействованные в формировании кратковременной памяти. Если воздействовать на окончание сенсорного нейрона (1), вводить в него циклический АМФ (2) или каталитическую единицу протеинкиназы А (3), это вызывает усиленное выделение нейромедиатора глутамата, что заставляет предположить, что все три вещества принимают участие в последовательности реакций, обеспечивающих кратковременную память.
Однако это еще ничего не говорило нам о том, как серотонин и циклический АМФ вызывают медленный синаптический потенциал и как этот потенциал связан с усилением выделения глутамата. В 1980 году в Париже, где я проводил серию семинаров в Коллеж де Франс, я познакомился со Стивеном Зигельбаумом. Стив был технически одаренным молодым биофизиком и специализировался на изучении свойств отдельных ионных каналов. Мы быстро нашли общий язык. Как выяснилось, судьбе было угодно, чтобы он незадолго до этого согласился перейти на работу на отделение фармакологии Колумбийского университета. Поэтому мы решили объединить усилия в Нью-Йорке и вместе исследовать биофизическую природу медленного синаптического потенциала.
Стив открыл одну из мишеней циклического АМФ и протеинкиназы А — калиевый ионный канал в мембране сенсорных нейронов, реагирующий на серотонин. Мы назвали его S-каналом, потому что он реагирует на серотонин и потому что его открыл Стив Зигельбаум. Этот канал открыт, когда нейрон находится в состоянии покоя, и принимает участие в поддержании потенциала покоя на его мембране. Стив выяснил, что канал присутствует в пресинаптических окончаниях и что можно вызывать его закрывание, либо воздействуя на клетку снаружи серотонином (первичным посредником), либо вводя внутрь клетки циклический АМФ (вторичный посредник) или протеинкиназу А. Закрывание калиевых каналов вызывает медленный синаптический потенциал, который когда-то и привлек наше внимание к циклическому АМФ.
Закрывание каналов также способствует усилению выделения глутамата. Когда эти каналы открыты, они вместе с другими калиевыми каналами принимают участие в поддержании мембранного потенциала покоя, а также в выходе калия из клетки во время реполяризации при потенциале действия. Но когда серотонин закрывает эти каналы, ионы калия выходят из клетки не так быстро, из-за чего продолжительность потенциала действия немного увеличивается за счет замедления реполяризации. Стив показал, что такое замедление потенциала действия дает кальцию больше времени на поступление в пресинаптические окончания, а кальций, как было показано Кацем в его экспериментах с гигантским синапсом кальмара, необходим для выделения глутамата. Кроме того, циклический АМФ и протеинкиназа А действуют непосредственно на аппарат, который обеспечивает слияние синаптических пузырьков с мембраной, тем самым еще больше увеличивая выделение глутамата.
К этим замечательным результатам, касающимся циклического АМФ, вскоре добавились важные результаты генетических исследований обучения у плодовых мух, ставших популярным подопытным животным за полвека с лишним до этого. В 1907 году Томас Морган, работавший в Колумбийском университете, начал использовать плодовую мушку дрозофилу в качестве модельного организма для генетических исследований — в связи с ее маленькими размерами и коротким репродуктивным циклом (двенадцать дней). Выбор оказался удачным, потому что у дрозофилы только четыре пары хромосом (у человека — двадцать три пары), благодаря чему проводить на ней генетические исследования сравнительно легко. Людям давно было очевидно, что многие физические характеристики животных (такие как форма тела, цвет глаз или скорость передвижения) наследуются. Но могут ли наследоваться психические характеристики, определяемые мозгом? Играют ли гены какую-то роль в психических явлениях, таких как память?
Первым, кто занялся изучением этого вопроса, используя современные методы, был Сеймур Бензер, сотрудник Калифорнийского технологического института. В 1967 году он начал серию блестящих экспериментов, в ходе которых дрозофил обрабатывали специальными веществами, вызывающими у них в отдельных генах случайные мутации, то есть наследственные изменения. Затем Бензер исследовал воздействие этих мутаций на обучение и память. Для изучения памяти у дрозофилы его ученики Чип Куинн и Ядин Дудай использовали классический метод выработки условного рефлекса. Они сажали мух в небольшую камеру и последовательно воздействовали на них двумя пахучими веществами. Затем в присутствии первого вещества на них действовали электрическими ударами, обучая избегать этого запаха. Потом мух помещали в другую камеру, в которой источники тех же двух запахов находились на противоположных концах. Обученные мухи избегали конца камеры, откуда исходил запах первого вещества, и устремлялись в другой, где находился источник второго запаха.
С помощью этого метода обучения Куинн и Дудай могли выявлять мух, лишенных способности запомнить, что запах первого вещества предвещает электрический удар. К 1974 году они испытали тысячи мух и выявили первого мутанта с нарушением кратковременной памяти. Бензер назвал этого мутанта dunce (тупица). В 1981 году Дункан Байерс, другой ученик Бензера, пойдя по стопам наших работ с аплизией, занялся изучением сигнального пути с участием циклического АМФ у мутантов dunce и обнаружил у них мутацию в гене, ответственном за утилизацию циклического АМФ. В результате в организме этих мутантов накапливается слишком много циклического АМФ, из-за чего, как можно предположить, их синапсы усиливаются до предела, что делает их нечувствительными к дальнейшим изменениям, и они не могут нормально функционировать. Впоследствии были выявлены и другие мутации в генах памяти. Они тоже оказались связаны с сигнальным путем с участием циклического АМФ.
Взаимодополняющие результаты, полученные в экспериментах с аплизией и дрозофилой (двумя очень непохожими подопытными животными, у которых исследовали разные формы обучения, используя разные подходы), были более чем обнадеживающими. Вместе они свидетельствовали, что механизмы, лежащие в основе простых форм имплицитной памяти, судя по всему, одинаковы у многих видов животных, в том числе у людей, причем для разных форм обучения, — в связи с тем, что эти механизмы эволюционно консервативны. Биохимия, а за ней и молекулярная биология, становилась мощными орудиями поиска общих черт биологического устройства разных организмов.
Эти открытия, сделанные на аплизии и дрозофиле, служили также дополнительным подтверждением важного биологического принципа: для создания новых адаптивных механизмов эволюции не требуется новых, специализированных молекул. Сигнальный путь с участием циклического АМФ задействован отнюдь не только в работе памяти. Как показал Сазерленд, он задействован даже не только в нейронах: и в кишечнике, и в почках, и в печени этот путь служит для обеспечения продолжительных метаболических изменений. Более того, из всех известных систем с использованием вторичных посредников система с циклическим АМФ, возможно, самая примитивная. Это важнейшая, а в некоторых случаях единственная система с использованием вторичных посредников, обнаруженная у одноклеточных организмов, таких как кишечная палочка, у которой циклический АМФ служит для сигнализации голода. Таким образом, биохимические процессы, лежащие в основе памяти, не были выработаны для выполнения именно этой функции. Наоборот, нейроны задействовали уже имеющуюся эффективную систему сигнализации, выполнявшую в других клетках иные функции, и стали использовать ее для обеспечения изменений синаптической силы, требуемых для формирования памяти.
Как отмечал специалист по молекулярной генетике Франсуа Жакоб, эволюция — не самобытный дизайнер, который ищет для новых проблем оригинальные решения. Эволюция — кустарь, работающий с подручными материалами. Она вновь и вновь немного по-разному использует одни и те же наборы генов. Она работает, варьируя существующие условия и просеивая случайные мутации в структуре генов, в результате которых возникают немного другие варианты белков и немного другие способы их использования в клетках. Большинство мутаций нейтрально или даже вредно и не выдерживает испытания временем. Только те редкие мутации, которые повышают шансы организма на выживание и размножение, с большой вероятностью сохраняются. Вот что пишет об этом Жакоб: «Работу естественного отбора нередко сравнивают с работой инженера. Однако это сравнение представляется неудачным. Во-первых <…> инженер работает в соответствии с заранее намеченным планом. Во-вторых, разрабатывая новую структуру, он не обязательно берет за основу какие-то старые структуры. Электрическая лампочка произошла не от свечки, а реактивный двигатель — не от двигателя внутреннего сгорания. <…> Наконец, новые объекты, выпускаемые инженером (по крайней мере, хорошим инженером), достигают пределов совершенства, которые возможны на основе технологий данного времени. Эволюция, в отличие от инженера, не делает ничего нового на пустом месте. Она работает с тем, что уже имеется, преобразуя ту или иную систему для выполнения новой функции или соединяя несколько систем, чтобы получить новую, более сложную. Если уж сравнивать с чем-то работу эволюции, придется признать, что она похожа на труд не столько инженера, сколько кустаря — bricolage, как мы говорим по-французски. Работа инженера зависит от того, имеются ли в его распоряжении сырье и инструменты, в точности соответствующие проекту, в то время как кустарь обходится чем придется. <…> Он пользуется всем, что окажется под рукой: старыми картонками, обрезками веревки, деревяшками и железками, — и делает из них какой-никакой рабочий предмет. Кустарь может подобрать какой-нибудь предмет, оказавшийся у него в запасе, и найти ему неожиданное применение. Из старого автомобильного колеса он может сделать вентилятор, а из сломанного столика — зонтик от солнца».
У живых организмов новые способности вырабатываются за счет незначительных изменений в молекулах уже имеющихся веществ и настройки их взаимодействия с другими имеющимися веществами. Психические процессы, свойственные человеку, долгое время считались не имеющими аналогов, поэтому некоторые ранние исследователи мозга ожидали, что найдут в глубинах нашего серого вещества много новых типов белков. На деле же наука обнаружила у нас в мозгу на удивление мало белков, не имеющих никаких аналогов, и не нашла ни одной уникальной для мозга системы сигнализации. Почти у всех белков мозга есть родственные им белки, выполняющие сходные функции в других клетках тела. Это относится даже к белкам, задействованным в процессах, которые происходят только в мозгу, например к белкам-рецепторам нейромедиаторов. Все живое, в том числе основание наших мыслей и воспоминаний, построено из одних и тех же структурных элементов.
Я подвел итоги первых сложившихся в систему открытий из области клеточной биологии кратковременной памяти в книге под названием «Клеточные основы поведения», опубликованной в 1976 году. В этой книге я подробно высказал (почти в форме манифеста), что для того, чтобы разобраться в поведении, необходимо применять радикальный редукционистский подход того же типа, что показал себя столь эффективным в других областях биологии. Примерно в то же время Стив Куффлер и Джон Николлс опубликовали книгу «От нейрона к мозгу», в которой подчеркивали огромные возможности клеточного подхода в нейробиологии. Пользуясь данными клеточной биологии, они рассказывали о том, как работают нейроны, как они образуют цепи в мозгу, а я — о связи поведения с мозгом. Стив тоже чувствовал эту связь и понимал, что нейробиология находится на пороге нового серьезного шага вперед.
Поэтому я был особенно рад, когда в августе 1980 года нам со Стивом представилась возможность совершить совместное путешествие. Нас обоих пригласили в Вену на церемонию приема в почетные члены Австрийского физиологического общества. Стив бежал из Вены в 1938 году. Сотрудникам медицинского факультета Венского университета нас представлял Вильгельм Ауэрсвальд — претенциозный профессор, мало сделавший в науке, который вел себя так, будто в причинах, вызвавших бегство двух сынов Вены из Австрии, не было ничего необычного. Он преспокойно сообщил, что Куффлер учился в Вене на медицинском факультете, а я жил на Северингассе — буквально в двух шагах от университета. Молчание, которым он обошел то, что на самом деле случилось с нами, говорило о многом. Ни Стив, ни я ничего не сказали в ответ на его слова.
Через два дня из Вены мы отправились на теплоходе вниз по Дунаю, в Будапешт, где приняли участие в Международном физиологическом конгрессе. Это была последняя серьезная конференция в жизни Стива. Он прочитал превосходный доклад. Вскоре после этого, в октябре 1980 года, он умер от сердечного приступа, случившегося после большого заплыва, в своем доме в Вудс-Хоул в штате Массачусетс, куда он приезжал на выходные.
Как и большинство коллег-нейробиологов, я был потрясен этой новостью. Мы все были многим обязаны Стиву и в чем-то зависели от него. Джек Макмахан, один из вернейших его учеников, описал то, что многие из нас почувствовали: «Как он мог с нами там поступить?»
В тот год я был председателем Нейробиологического общества и возглавлял оргкомитет, отвечавший за ежегодный ноябрьский симпозиум. Он проходил в Лос-Анджелесе всего через несколько недель после смерти Стива и на него собралось около десяти тысяч нейробиологов. Дэвид Хьюбел прочитал на этом симпозиуме замечательный доклад памяти Стива. Он показывал слайды и рассказывал о том, каким глубоким, проницательным и великодушным человеком был Стив, как много он значил для всех нас. Думаю, никто из наших американских коллег с тех пор не имел такого влияния и не снискал такой всеобщей любви, как Стив. Джек Макмахан организовал издание книги в память о нем. Я писал в ней: «Готовя этот раздел, я чувствую, что Стив во многом по-прежнему с нами. После Олдена Спенсера у меня нет другого коллеги, о котором я думал бы больше, чем о нем, и которого мне бы так же не хватало».
Смерть Стива Куффлера означала конец целой эпохи — эпохи, когда сообщество нейробиологов было еще сравнительно небольшим и его деятельность сосредоточивалась на клетке как единице устройства мозга. Смерть Стива совпала со слиянием нейробиологии с молекулярной биологией — событием, которое резко расширило область приложения нейробиологии и увеличило число ученых, работающих в этой области. Изменения отразились и на моей работе: в 1980 году мои клеточные и биохимические исследования обучения и памяти были уже близки к завершению. К тому времени мне стало ясно, что повышение концентрации циклического АМФ и усиление выделения медиатора, вызываемые серотонином в ответ на единственное обучающее событие, длятся лишь минуты. В продолжительном усилении связей, которое длится дни и недели, должно быть задействовано что-то еще — возможно, изменения в экспрессии генов, а также анатомические изменения. Поэтому я обратился к изучению генов.
Я был готов к этому. Долговременная память уже начала занимать мое воображение. Как человеку удается на всю жизнь запоминать события детства? Мать Дениз, Сара Бистрен, от которой Дениз и ее брат Жан-Клод, а также их супруги И дети унаследовали вкус к декоративному искусству (мебели, вазам и лампам в стиле ар-нуво), редко говорила со мной о моей научной работе. Но, должно быть, она как-то почувствовала, что я готов был заняться генами и долговременной памятью.
На мой день рождения 7 ноября 1979 года, когда мне исполнилось пятьдесят, она купила мне прекрасную вазу в венском стиле, изготовленную в городе Теплице (рис. 16–5), и подарила ее вместе с надписью следующего содержания:
«Дорогой Эрик!Эта ваза из Теплице, вид Венского леса, ностальгия, которую излучают деревья, цветы, свет, закат, принесут тебе память былых времен — воспоминания о твоем детстве. И когда ты будешь пробегать вдоль леса в Ривердейле, ностальгия по венскому лесу охватит тебя и ненадолго заставит забыть о событиях твоей повседневной жизни.С любовью, Сара»
Так Сара Бистрен сформулировала мою задачу.

16–5. Ваза из города Теплице (Из архива Эрика Канделя).

