Книга: Эволюция. Классические идеи в свете новых открытий
Назад: Избыточность и эволюционная пластичность
Дальше: Маски и их прототипы в пьесе EvoDevo
Чтобы построить новое, нужно расшатать старое
Простейшие эволюционные модели предполагают, что новые адаптивные признаки развиваются по следующей схеме.
• Сначала происходит случайная мутация в ДНК.
• Эта мутация неким вполне определенным образом проявляется в фенотипе, т. е. меняет строение, физиологию или поведение организма.
• Если изменение фенотипа повысило приспособленность организма (т. е. увеличилась эффективность передачи генов следующим поколениям), то с течением времени частота встречаемости мутантного гена в популяции будет расти. Этот автоматический и неизбежный процесс известен под названием естественного отбора.
• В конце концов мутация может зафиксироваться. Это значит, что частота мутантного гена в популяции достигла 100 % и новый признак стал нормой для всех особей.
Нет сомнений, что часто все именно так и происходит. Многие рассмотренные нами примеры появления эволюционных новшеств отлично описываются этой классической схемой.
Однако данная модель основана на допущении, которое не всегда справедливо. Речь идет о том пункте, где предполагается, что фенотипическое проявление мутации является «вполне определенным». Генетики прекрасно знают, что реальные взаимосвязи между генами и признаками часто не столь однозначны. Гены вообще определяют фенотип не строго, а вероятностным образом. В общем случае мутация приводит не к какому-то строго определенному изменению фенотипа, а к изменению вероятностей реализации тех или иных фенотипов, причем эти вероятности зависят не только от самого мутантного гена, но и от «генетического контекста», т. е. от других генов генома. Один и тот же мутантный ген у одних особей может привести к резко измененному фенотипу, у других — не дать видимого эффекта (даже в гомозиготном состоянии), так что фенотип получится «нормальный». Как мы уже знаем, это называют неполной пенетрантностью мутаций.
Для нормального развития необходима слаженная работа всех генов, поэтому, строго говоря, фенотипический признак не может определяться только одним геном — в конечном счете он зависит от всех генов генома. Поэтому обычно между генами и признаками нет ничего похожего на связь «один к одному».
Изменение, произошедшее на уровне ДНК (мутация), влияет не непосредственно на фенотипический признак, а на ход индивидуального развития. Мутация может отклонить ход развития в ту или иную сторону. Однако онтогенез обычно обладает высокой помехоустойчивостью. В ходе эволюции отбор закрепляет такие генетические изменения, которые повышают способность онтогенеза противостоять нарушениям, компенсировать помехи и возвращаться на «магистральный путь». Эту способность онтогенеза невзирая на помехи приходить к одному и тому же итогу («нормальному» фенотипу) называют эквифинальностьюонтогенеза.
Низкая пенетрантность многих вредных мутаций — яркое проявление помехоустойчивости онтогенеза. Во многих случаях растущему организму удается справиться с мутациями, которые нарушают нормальный ход развития. Например, мутантные белки, синтезированные на основе мутантных генов, могут тем не менее нормально функционировать, если шапероны «насильно» придадут им правильную трехмерную конфигурацию.
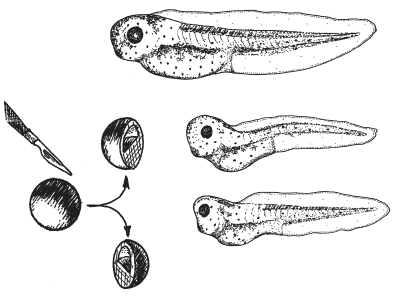 Пример эквифинальности. Если аккуратно разрезать пополам скальпелем ранний эмбрион (бластулу) шпорцевой ляшушкиXenopus , то из каждой половинки разовьется не полголовастика, как можно было бы предположить, а нормальный целый головастик. Нужно только, чтобы в каждую половинку попало по кусочку организатора Шпемана. Как мудро заметил обнаруживший этот факт эмбриолог Эдди Де Робертис из Калифорнийского университета в Лос-Анджелесе, естественный отбор не стал бы создавать такую саморегуляцию только на случай, что придет любознательный эмбриолог со скальпелем и начнет резать бластулы пополам (De Robertis, 2009 ). Разумеется, это результат отбора на стабильность онтогенеза, на его способность противостоять любым помехам: внутренне-стохастическим, мутационным или средовым.
Пример эквифинальности. Если аккуратно разрезать пополам скальпелем ранний эмбрион (бластулу) шпорцевой ляшушкиXenopus , то из каждой половинки разовьется не полголовастика, как можно было бы предположить, а нормальный целый головастик. Нужно только, чтобы в каждую половинку попало по кусочку организатора Шпемана. Как мудро заметил обнаруживший этот факт эмбриолог Эдди Де Робертис из Калифорнийского университета в Лос-Анджелесе, естественный отбор не стал бы создавать такую саморегуляцию только на случай, что придет любознательный эмбриолог со скальпелем и начнет резать бластулы пополам (De Robertis, 2009 ). Разумеется, это результат отбора на стабильность онтогенеза, на его способность противостоять любым помехам: внутренне-стохастическим, мутационным или средовым.
Кроме генов, на ход развития (и соответственно, на итоговый фенотип) влияют и другие факторы: например, температура и химический состав среды, в которой происходит развитие. Самое интересное, что влияние мутаций и изменений внешних условий может быть очень похожим. Одно и то же изменение фенотипа (например, какое-нибудь уродство) можно получить и в результате мутации, и в результате физического воздействия на развивающийся организм. Например, подвергая личинок дрозофилы тепловому шоку, можно получить такие же уродства, как и те, что возникают в результате мутаций. И наоборот: если взять чуть ли не любое уродство, появляющееся в результате перегрева, то в большинстве случаев оказывается, что некоторые мутации могут приводить к такому же уродству без всякого перегрева (или при менее интенсивном перегреве). Это явление называется генокопированием модификаций. Можно сказать, что у организма уже имеется потенциально возможный альтернативный путь развития, который приводит к появлению нового признака (даже если в нормальных условиях этот путь никогда не реализуется). Нужно лишь подобрать условия — внешние (температуру) или внутренние (мутацию), — в которых развитие пойдет по этому альтернативному пути.
Отбор обычно способствует повышению роли внутренних (генетических) и снижению роли внешних регуляторов онтогенеза. Онтогенез под действием отбора стабилизируется и приобретает все большую автономность, становится все более помехоустойчивым. Это значит, что геном постепенно меняется таким образом, чтобы обеспечивать реализацию «нормального» фенотипа со все большей вероятностью.
На основе подобных фактов и рассуждений некоторые исследователи, среди которых следует упомянуть И. И. Шмальгаузена, К. Х. Уоддингтона (1905–1975) и М. А. Шишкина (р. 1936), пришли к выводу, что именно онтогенез (как сложная помехоустойчивая система индивидуального развития) является главным действующим лицом эволюционной драмы. В наиболее абсолютизированной форме эти воззрения отстаивает М. А. Шишкин, который считает, что основной механизм формирования эволюционных новшеств существенно отличается от приведенного выше и выглядит примерно так ( Шишкин, 1988, 2006).
• Сильное внешнее воздействие (например, резкое изменение среды) приводит к дестабилизации онтогенеза.
• Это автоматически ведет к появлению разнообразных аномальных фенотипов (реализуются доселе скрытые альтернативные пути развития).
• Если какие-то из аномальных фенотипов окажутся «удачными» (адаптивными в новых условиях), отбор в дальнейшем будет закреплять такие мутации, которые повысят вероятность реализации именно этого альтернативного пути развития. В результате аномалия постепенно станет новой нормой.
• В ходе стабилизации новой нормы возникнут новые скрытые альтернативные пути развития, которые смогут реализоваться при следующем «кризисе».
Главная особенность этой модели в том, что эволюционное преобразование не начинается с генетических изменений, а заканчивается ими. Новый фенотип сначала появляется как редкая аномалия ( морфоз) — отклонение онтогенеза от нормального пути при неизменном геноме. В дальнейшем отбор постепенно вписываетновый путь онтогенеза в геном, фиксирует его на генетическом уровне, т. е. делает его все более генетически детерминированным, стабильным и помехоустойчивым. По мнению М. А. Шишкина, «эволюционные изменения начинаются с фенотипа и распространяются по мере их стабилизации в направлении генома, а не наоборот». Главное — не путать эти взгляды с ламаркизмом и помнить, что фиксация эволюционных изменений в геноме происходит на основе «дарвиновского» механизма, т. е. путем закрепления естественным отбором случайных мутаций.
Впрочем, ясно, что первоначальное «возмущающее воздействие» в этой схеме не обязательно должно быть внешним — это может быть и мутация, но не такая, которая сразу приводит к появлению нового стабильного признака, а такая, которая вносит хаос в систему индивидуального развития (дестабилизирует онтогенез). В этом случае отличие модели Шишкина от классической становится менее контрастным.
Чего этой теории сильно не хватает, так это хороших иллюстраций, т. е. детальных исследований (в том числе молекулярно-генетических), показывающих реальность такого эволюционного механизма. Исследование американских микробиологов, выполненное в 2009 году на почвенной бактерии Bacillus subtilis, отчасти восполняет этот пробел ( Eldar et al., 2009). Разумеется, авторы пользуются другой терминологией и не цитируют ни Шмальгаузена, ни Шишкина, хотя, надо отдать им должное, Уоддингтона они все-таки упомянули.
Бактерия Bacillus subtilis— излюбленный модельный объект генетиков и молекулярных биологов. О повадках этого микроба мы немного рассказали в книге «Рождение сложности». При наступлении неблагоприятных условий бактерии переходят к каннибализму, причем у некоторых особей (вот она, клеточная стохастика!) отключаются защитные механизмы, что позволяет другим их съесть. Если это не помогает и голод продолжается, бактерии производят споры, способные переждать тяжелые времена. Процесс образования спор (споруляция) у B. subtilisподробно изучен.
Исследователи работали с 53 мутантными штаммами B. subtilis. Штаммы были сконструированы при помощи генно-инженерных методов. В геном бактерии вносились различные мутации, нарушающие работу гена spoIIR. Этот ген необходим для того, чтобы формирующаяся спора могла передать в материнскую клетку сигнал, блокирующий формирование второй перегородки-септы (см. рисунок).
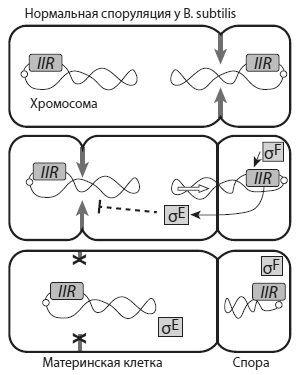 Схема споруляции уB. subtilis .Серыми стрелками показано образование септы (перегородки), которая отделяет будущую спору от материнской клетки. Кольцевые хромосомы показаны в виде двух перекрученных петель. Регуляторный белокσ F , образующийся в будущей споре, активизирует ген spoIIR, который на рисунке для краткости обозначен IIR. Это приводит к тому, что другой сигнальный белок,σ Е , блокирует формирование второй септы на противоположном конце материнской клетки. ИзEldar et al., 2009 .
Схема споруляции уB. subtilis .Серыми стрелками показано образование септы (перегородки), которая отделяет будущую спору от материнской клетки. Кольцевые хромосомы показаны в виде двух перекрученных петель. Регуляторный белокσ F , образующийся в будущей споре, активизирует ген spoIIR, который на рисунке для краткости обозначен IIR. Это приводит к тому, что другой сигнальный белок,σ Е , блокирует формирование второй септы на противоположном конце материнской клетки. ИзEldar et al., 2009 .
Хотя мутации были разные, фенотипический эффект у них оказался похожим. Во всех случаях произошла дестабилизациясистемы спорообразования. Вместо одного-единственного пути развития, который характерен для «диких» бактерий (образование одной споры в каждой клетке), бактерии-мутанты демонстрировали несколько вариантов развития, причем выбор того или иного варианта осуществлялся случайным образом. Генотип влиял только на вероятность, т. е. частоту реализации каждого из них. Варианты были следующие.
1. Нормальная споруляция. В этом случае, несмотря на мутацию, «пытающуюся» нарушить нормальный ход развития, белок σ Eвсе равно образуется в материнской клетке в нужное время и в достаточном количестве. Поэтому образование второй септы своевременно блокируется, и дальнейшее развитие идет так же, как у «диких» бактерий. Это можно считать проявлением помехоустойчивостионтогенеза, а можно назвать неполной пенетрантностьювредной мутации, что по сути одно и то же.
2. Неудачная попытка образовать сразу две споры в одной материнской клетке. Белок σ Eвырабатывается в недостаточном количестве (или слишком поздно), и в результате вторая септа успевает сформироваться. Получается нежизнеспособная конструкция из двух незрелых спор, содержащих по хромосоме, а между ними — лишенная хромосомы материнская клетка. Это заканчивается гибелью всех троих.
3. Прерванная споруляция. Начавшийся процесс формирования споры прерывается, «зачаток» споры отмирает, а материнская клетка начинает расти. При этом в ней происходит репликация ДНК, т. е. хромосома сначала удваивается, а потом иногда и утраивается. Такая клетка может затем перейти к обычному делению, а может «вспомнить», что собиралась заняться производством спор, и тогда начинается самое интересное: возникает вариант 4.
4. Споры-близнецы. Если в клетке, прервавшей споруляцию, произойдет утроение хромосомы, а потом споруляция возобновится, то получается материнская клетка (со своей хромосомой), на концах которой образуются две жизнеспособные споры, тоже содержащие каждая по одной хромосоме. Эти споры в подходящих условиях нормально прорастают (т. е. превращаются в обычных питающихся и делящихся бактерий) и вообще ничем не отличаются от нормальных спор B. subtilis.
5. Одиночная спора в диплоидной материнской клетке. Клетка с тремя хромосомами может произвести не две, а одну спору. В этом случае в материнской клетке остается две хромосомы. Спора при этом тоже получается нормальная, как и в варианте 4.
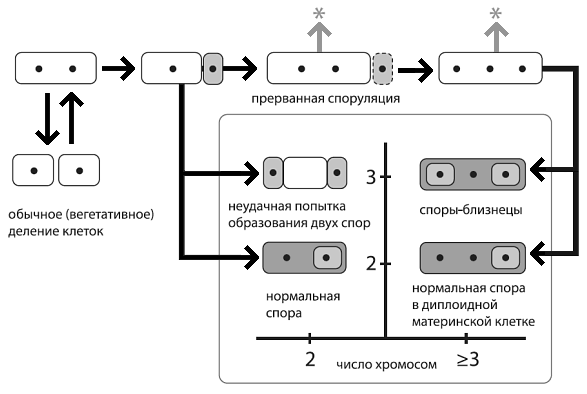 Альтернативные пути развития у бактерий с дестабилизированной системой споруляции.Черными точками показаны хромосомы.Светло-серым цветом выделены формирующиеся споры (в них вырабатывается белокσF ),темно-серым — материнские клетки (в них вырабатывается белокσE ). По горизонтальной оси — число хромосом, по вертикальной — число «отсеков», на которые разделилась исходная клетка.Стрелка со звездочкой символизирует возможность возвращения к обычному размножению путем деления. ИзEldar et al., 2009 .
Альтернативные пути развития у бактерий с дестабилизированной системой споруляции.Черными точками показаны хромосомы.Светло-серым цветом выделены формирующиеся споры (в них вырабатывается белокσF ),темно-серым — материнские клетки (в них вырабатывается белокσE ). По горизонтальной оси — число хромосом, по вертикальной — число «отсеков», на которые разделилась исходная клетка.Стрелка со звездочкой символизирует возможность возвращения к обычному размножению путем деления. ИзEldar et al., 2009 .
Таким образом, дестабилизация развития приводит к тому, что наряду с «нормой» (вариант 1) появляется четыре «морфоза», или альтернативных пути развития (варианты 2–5). Весь этот спектр морфозов проявляется в генетически однородных популяциях каждого из 53 мутантных штаммов. Иными словами, мутантные бактерии с одинаковым геномом «выбирают» один из пяти путей развития случайным образом. От конкретной мутации — т. е. от того, как именно нарушена работа гена spoIIR, — зависит лишь частота реализации этих путей.
Из четырех возникших морфозов один безусловно вреден (№ 2), два других (№ 3 и 5) не создают ничего принципиально нового и вряд ли могут оказаться полезными — скорее всего, они приведут только к лишним тратам ресурсов, т. е. окажутся неадаптивными.
Наиболее интересен морфоз № 4 — образование жизнеспособных спор-близнецов. В принципе такое изменение механизма споруляции может оказаться полезным — например, в том случае, если условия среды очень нестабильны, обычный способ размножения (деление клеток надвое) реализовать трудно и удобнее размножаться спорами.
Если формирование спор-близнецов может быть полезным, то следует ожидать, что у каких-нибудь бактерий такой способ размножения закрепился и стал нормой. Это действительно так: у многих бактерий из группы клостридий «близнецовая споруляция» является нормальным способом размножения. Авторы изучили близнецовую споруляцию у некоторых клостридий и пришли к выводу, что она протекает точно так же, как у мутантных B. subtilis. Надо сказать, что генные системы, управляющие споруляцией у клостридий и B. subtilis, гомологичны, т. е. имеют единое происхождение, однако в природе у B. subtilisблизнецовая споруляция никогда не встречается.
Итак, дестабилизация онтогенеза привела к появлению морфоза (аномального пути развития), который потенциально может оказаться полезным. Однако в исследованных мутантных штаммах этот путь развития реализуется лишь у небольшой части особей, т. е. представляет собой редкую аномалию. Сначала клетка должна «выбрать» вариант развития № 3 («прерванная споруляция»); частота такого выбора зависит от конкретной мутации. Из особей, сделавших такой выбор, примерно 25 % снова приступают к споруляции, а из них лишь у 5 % образуются споры-близнецы. Таким образом, потенциально полезный морфоз возникает у мутантов очень редко. Может ли он в дальнейшем стабилизироваться, иными словами — стать нормой?
Первопричиной этого морфоза, как и трех других, является изменение активности гена spoIIR. Выяснилось, однако, что вероятность выбора клетками одного из возможных путей развития лишь на 15 % определяется силой мутации (т. е. тем, насколько изменилась экспрессия spoIIR). Остальные 85 % вариабельности клеток по этому признаку зависят от случайности. Регулируя только лишь экспрессию spoIIR, невозможно добиться стабилизации одного из морфозов. Даже повлиять на частоту его реализации можно лишь в очень узких пределах. Иными словами, такие мутации не могут отклонить путь развития в каком-то конкретном направлении — они могут только дестабилизировать систему, т. е. внести элемент хаоса в программу развития.
Следовательно, для того чтобы полезный морфоз стабилизировался и стал «нормой», необходимы дополнительные мутации. Какие именно и существуют ли они в природе? Чтобы ответить на этот вопрос, авторам пришлось изучить взаимовлияние двух процессов, играющих ключевую роль в споруляции. Первый процесс — это репликация ДНК, в ходе которой в клетке увеличивается число хромосом; второй — образование септ (перегородок, отделяющих будущую спору от материнской клетки).
Как выяснилось, вероятность формирования спор-близнецов у бактерий с дестабилизированным онтогенезом зависит от комбинации двух факторов: скорости формирования септ и скорости репликации. Для успешного образования спор-близнецов необходимо, чтобы в материнской клетке сначалаобразовалась дополнительная хромосома и только потомначала расти вторая септа. Если вторая септа образуется в материнской клетке с одной хромосомой, развитие пойдет по пути № 2 и закончится гибелью. Если же вторая септа начнет образовываться в материнской клетке с двумя хромосомами, получаются споры-близнецы.
Исследователи предположили, что можно увеличить вероятность образования спор-близнецов, повысив у мутантных бактерий темп репликации. Это предположение блестяще подтвердилось. Были испытаны две разные мутации, ускоряющие репликацию. В результате процент клеток, производящих по две жизнеспособные споры, вырос от долей процента до 30 %, что сопоставимо с естественной частотой «близнецовой споруляции» у клостридий. Более того, ускорение репликации у мутантных B. subtilisпривело к тому, что стал излишним выбор варианта № 3 (прерванная споруляция): клетки смогли осуществлять близнецовую споруляцию напрямую, минуя этап прерванной споруляции. Авторы также попробовали внести мутацию, ускоряющую репликацию, в геном «диких» B. subtilis. Это привело к тому, что у них иногда — в виде редкой аномалии — стали образовываться споры-близнецы.
По-видимому, дестабилизация развития может существенно облегчать формирование новых адаптаций. Дестабилизация как бы перекидывает мостик от одного стабильного состояния к другому, что особенно ценно в том случае, когда этот переход нельзя осуществить за счет одной-единственной мутации (перепрыгнуть пропасть одним прыжком).
Действительно, чтобы бактерии, практикующие одиночную споруляцию, как B. subtilis, начали систематически осуществлять близнецовую споруляцию, им недостаточно приобрести какую-то одну мутацию, которая сразу переведет их из одного устойчивого состояния в другое. Для этого нужны как минимум две мутации, одна из которых повысит темп образования септ (именно такой эффект дает изменение активности spoIIR), а другая увеличит скорость репликации, чтобы появление второй септы вело не к гибели, а к созданию двух жизнеспособных спор. Но две мутации не могут, как по заказу, появиться одновременно. Тут-то и приходит на помощь механизм дестабилизации онтогенеза. Первая мутация позволяет бактериям реализовать один из «скрытых» путей развития — пусть и с очень низкой частотой. Если данный морфоз окажется выгодным, это даст шанс дестабилизированной популяции продержаться некоторое время, пока у какой-нибудь бактерии не возникнет вторая мутация, повышающая скорость репликации. Это приведет к тому, что полезная аномалия закрепится — станет новой нормой. В эволюции клостридий такое, по-видимому, происходило не раз.
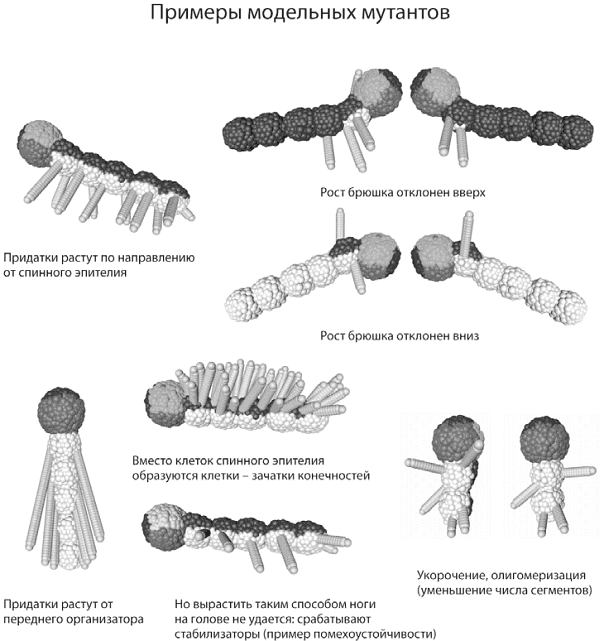 Примеры интересных новых фенотипов, возникающих при внесении в генотип нашего модельного сегментированного червячка единичных случайных изменений — мутаций. Важным побочным эффектом «зарегулированности» онтогенеза является неожиданномалое число возможных изменений фенотипа, возникающих в результате единичных мутаций. Многие разные мутации приводят к одному и тому же фенотипическому эффекту, и в целом у фенотипа оказывается гораздо меньше «степеней свободы», чем у генотипа. Иными словами, если генетическая программа развития достаточно сильно стабилизирована обратными связями и «избыточными» регуляторами, число возможных путей эволюционных изменений фенотипа резко сокращается. Эволюция становится канализированной, т. е. во многом предопределенной свойствами генетической программы онтогенеза.
Примеры интересных новых фенотипов, возникающих при внесении в генотип нашего модельного сегментированного червячка единичных случайных изменений — мутаций. Важным побочным эффектом «зарегулированности» онтогенеза является неожиданномалое число возможных изменений фенотипа, возникающих в результате единичных мутаций. Многие разные мутации приводят к одному и тому же фенотипическому эффекту, и в целом у фенотипа оказывается гораздо меньше «степеней свободы», чем у генотипа. Иными словами, если генетическая программа развития достаточно сильно стабилизирована обратными связями и «избыточными» регуляторами, число возможных путей эволюционных изменений фенотипа резко сокращается. Эволюция становится канализированной, т. е. во многом предопределенной свойствами генетической программы онтогенеза.
Это исследование показало реалистичность сценария, предлагаемого М. А. Шишкиным и его единомышленниками. Имеет ли смысл абсолютизировать этот механизм (т. е. утверждать, что путь к новым адаптациям всегдалежит через дестабилизацию онтогенеза и последующую генетическую фиксацию морфозов) — вопрос отдельный, и мы склоняемся к отрицательному ответу. Но все же следует учитывать, что каждый онтогенетический путь может быть пройден несколькими дорожками, явными или тайными, и эти дорожки предопределены структурой и внутренней логикой устройства как организма, так и его онтогенеза. И если смотреть на новые признаки как на результат проявления скрытых путей развития, то становится легче понять, почему прогрессивная эволюция на нашей планете в целом идет довольно бодрыми темпами. По-видимому, мутация, вмешивающаяся в развертывание сложной, устойчивой и саморегулирующейся программы развития, с большей вероятностью породит что-то новое и осмысленное, чем мутация, непосредственно и однозначно влияющая на конечный результат этого развития. Внеся случайное изменение в готовый осмысленный текст, мы почти наверняка его испортим. Но если у нас есть хорошая, умнаяпрограмма генерации осмысленных текстов и мы вмешаемся в ее работу на каких-то относительно ранних этапах, то появится ненулевой шанс получить на выходе что-то интересное.
Многочисленные обратные связи и стабилизирующие контуры, которыми переполнены (в результате действия отбора «на стабильность») генетические «программы» онтогенеза, как раз и придают им свойство, которое компьютерщики неформально называют умом. «Умная» программа способна справиться с разными нештатными ситуациями, в том числе даже такими, которые не были предусмотрены программистом (или естественным отбором, если мы говорим об онтогенезе реальных организмов).
Более того, даже если «умная» программа не сможет полностью скомпенсировать помехи и выдать абсолютно нормальный результат, тот искаженныйрезультат, который она в итоге выдаст, имеет хороший шанс оказаться интересным, т. е. эволюционно перспективным, адаптивным в каких-то условиях. В этом, возможно, одна из причин самоускоряющегося усложнения, повышения «уровня организации», которое мы наблюдаем в некоторых эволюционных линиях. В таких, например, как животные в целом и позвоночные в частности.

