Книга: Эволюция. Классические идеи в свете новых открытий
Назад: Глава 8 Эволюция алгоритмов
Дальше: Неустранимая случайность
Чудо самоорганизации
Говорят, что курица — всего лишь способ, посредством которого яйцо производит другое яйцо. Эта шутка подчеркивает однобокость такого взгляда на живое существо, при котором какая-то произвольно выбранная стадия его развития (например, курица) считается «главной», а другие (например, яйцо) — «второстепенными».
Эволюционируют не взрослые организмы. Эволюционируют не ювенильные (незрелые) организмы. Эволюционирует вся система целиком, система продвижения от яйца до половозрелого организма, т. е. онтогенез. Иными словами, эволюционируют онтогенезы.
Онтогенезы основаны на самоорганизации. Нельзя понять эволюцию, не понимая, что такое самоорганизация и как она работает. К сожалению, понять это не очень просто, потому что наши мозги плохо приспособлены для этого. Но мы все же постараемся.
Всем хорошо знаком такой наглядный пример самоорганизации, как образование морозных узоров на стекле. При этом у нас на глазах из «простого» (из хаотического движения молекул водяного пара) рождается «сложное» — изысканные ледяные картины. Мы знаем, что эти узоры никто не рисует, они образуются сами. Мы не считаем это чудом (хотя это очень похоже на чудо!) и не привлекаем для объяснения сверхъестественные силы (хотя и говорим полушутя, что это художества Деда Мороза).
То же и с развитием многоклеточного организма. Оно еще больше похоже на чудо. Из комочка клеток, возникших в результате первых делений зиготы, — одинаковых клеток, у которых нет ни целей, ни планов, ни разума, — сам собой, словно по мановению волшебной палочки, развивается сложнейший организм.
Как это получается? Как получается, что клетки зародыша, исходно одинаковые, в какой-то момент вдруг «понимают», что одни должны стать печенью, другие — костями, третьи — мозгами? Казалось бы, должен быть какой-то «центральный управляющий орган». Где же он?
Удивительно, но никакого управляющего центра нет. Множество одинаковых объектов — например, делящихся клеток — могут формировать сложные структуры, создавая видимость осмысленного поведения, без всякого централизованного руководства. Это и есть самоорганизация (или самосборка). И если она вызывает у нас ощущение чуда, то это проблема нашей психики, а не объективной реальности.
—————ПредопределенностьОднажды великий биолог Джон Холдейн (1892–1964), известный своими афоризмами, читал публичную лекцию, после которой одна дама встала и сказала: «А все-таки я не могу поверить, что из одноклеточного организма само собой могло развиться такое сложное и совершенное существо, как человек, даже если у него были на это миллиарды лет». Холдейн ответил: «Мадам, но вы сами это проделали. И у вас ушло на это всего девять месяцев».Конечно, мы могли бы на это возразить, что между онтогенезом и эволюцией есть принципиальная разница: в случае онтогенеза исходная клетка (зигота) содержит некую генетическую «программу развития», сложившуюся в ходе предшествующей эволюции. В случае эволюционного развития аналогичной программы нет.Но если подумать, это различие хоть и существует, оно, возможно, не настолько глубоко, как кажется на первый взгляд. Ведь на самом деле в зиготе нет «программы развития» в чистом виде. Если рассматривать геном как программу, то это не программа развития эмбриона. Это программа поведения клетки— одной клетки, и не более того.Самосборка организма из множества делящихся клеток с одним и тем же геномом — процесс относительно жестко детерминированный, и детерминирован он как раз генетической программой поведения клетки. На генетическую программу накладываются модифицирующие или корректирующие ее факторы среды. Но ведь и эволюция тоже процесс отчасти детерминированный и во многом зависящий от генома эволюционирующих организмов. Может быть, эволюцию тоже можно рассматривать как своего рода самосборку, ход которой предопределен, с одной стороны, геномом эволюционирующего организма, с другой — факторами среды.Наследственная изменчивость и дифференциальное размножение — это механизм самоорганизации, благодаря которому из простейших существ сами собой с неизбежностью развиваются те самые endless forms most beautiful and most wonderful(«бесчисленные прекрасные и удивительные формы»), о которых с восхищением писал Дарвин в заключительном разделе «Происхождения видов». Все это поразительное многообразие жизни было в каком-то смысле «закодировано» в самых первых живых существах, в их ДНК или РНК, и в условиях среды, где им предстояло эволюционировать. Конечно, с учетом того, что условия среды будут многократно изменены самими эволюционирующими существами.—————
Принципы онтогенеза (индивидуального развития) многоклеточных контринтуитивны, трудны для понимания, потому что мозг Homo sapiensплохо приспособлен для понимания процессов самоорганизации. Наше мышление адаптировано для постановки целей и поиска путей их достижения. Мы должны все контролировать (или понимать, кто контролирует и почему), иначе ничего не будет, кроме хаоса (отсюда, кстати, и живучесть креационизма, и конспирологические мифы о всесильных «мировых закулисах»).
Поэтому когда мы видим, как из чего-то простого само собой вдруг образуется что-то сложное, нам это кажется чудом, и мы начинаем подозревать воздействие какой-то особой организующей силы. Мы начинаем изобретать лишние сущности. В биологии таких сущностей напридумано множество: от вмешательства инопланетян и Божественного сотворения до ламарковского стремления к совершенству, радиальной энергии Тейяра де Шардена, активности Ю. В. Чайковского и т. д.
Ну а в данном случае, который показан на рисунке, — в случае самосборки снежинки из хаотически движущихся молекул воды — совершенно очевидно вмешательство Деда Мороза.
Где закодировано строение снежинки? Пожалуй, мы не сильно ошибемся, если скажем, что оно закодировано в свойствах молекулы воды в таком же смысле, в каком взрослый фенотип закодирован в геноме зиготы. И внешняя среда в обоих случаях сильно влияет на результат.
Контринтуитивность онтогенеза (как и эволюции) порождает множество проблем, и многие теоретики действительно на этом спотыкались. Попробуем разобраться постепенно.
Рассмотрим простой комочек клеток — ранний зародыш, который образовался из яйцеклетки в результате нескольких первых делений. Каждая клетка зародыша имеет один и тот же геном. Геном определяет свойства клетки, это ее «программа поведения». Программа у всех клеток зародыша одинаковая. Однако клетки начинают вести себя по-разному: одни превращаются в клетки кожи, другие — в клетки кишечника, третьи дают начало нервной системе.
Это происходит благодаря тому, что клетки обмениваются информацией — посылают друг другу химические сигналы и меняют свое поведение в зависимости от того, какие сигналы они получили от соседей. Кое-какие сигналы приходят и из внешнего мира. Например, клетки зародышей растений чувствуют земное притяжение и принимают его в расчет, когда решают, как им себя вести. Наконец, яйцеклетка с самого начала имеет простенькую разметку: один ее полюс отличается от другого по концентрации целого ряда веществ. Реальные яйцеклетки животных поляризованы, в частности, за счет того, что на одной их стороне содержится больше запасных питательных веществ (желтка), чем на другой.
Программа поведения у всех клеток зародыша изначально одна и та же, но она может состоять из нескольких отдельных наборов правил. То, какой из них данная клетка будет выполнять, зависит от получаемых клеткой сигналов. Каждое отдельное правило выглядит примерно так: если выполняются такие-то условия, сделай такое-то действие. Основное действие клетки — включение или выключение определенных генов. Это меняет свойства клетки, она начинает по-другому себя вести, по-другому реагировать на сигналы.
Правила поведения клетки определяются системой взаимодействий между генами — генно-регуляторными сетями. На физическом уровне правила «сделаны» из рецепторов, транскрипционных факторов, энхансеров, сигнальных молекул и белков, осуществляющих синтез и транспортировку этих молекул. Например, правило «Если получен сигнал А, начни выделять вещество Б», может быть сделано из рецептора вещества А, который активирует транскрипционный фактор В, который прикрепляется к энхансеру Г, расположенному около гена Д, который кодирует фермент, отвечающий за синтез вещества Б.
Сигнальные вещества, на которые клетки эмбриона реагируют, меняя свое поведение в зависимости от их концентрации, называются морфогенами. В приведенном примере в роли морфогена выступает вещество А.
Но как же все-таки получается, что клетки зародыша, имеющие одинаковую программу поведения и находящиеся, казалось бы, в одинаковых условиях, тем не менее ведут себя по-разному? Дело в том, что на самом деле они находятся в разныхусловиях. Так получается само собой в процессе деления клеток. Кто-то оказался внутри, кто-то снаружи, кто-то снизу, кто-то сверху, в ком-то концентрация морфогена А выше (потому что данная клетка сформировалась из той части яйцеклетки, где этого вещества было много), в ком-то ниже.
Еще у клеток может быть «счетчик делений», который сообщает им, сколько раз яйцеклетка уже поделилась. Этот счетчик тоже химический: в яйцеклетке изначально были определенные вещества (например, молекулы матричных РНК, считанные еще с генов материнского организма), запас которых не пополняется во время развития зародыша, и по тому, сколько в клетке осталось этих веществ, можно понять, сколько делений прошло с момента начала развития.
Программа поведения клетки может содержать, например, такие команды:
«Если ты снаружи от других клеток, и если концентрация морфогена А такая-то, и если концентрация морфогена Б равна нулю, и если с момента начала развития пройдено десять делений, то начни выделять морфоген Б».
К чему приведет выполнение такой инструкции? Оно приведет к тому, что на поверхности зародыша в определенный момент (после десяти делений) появится одна-единственная клетка, выделяющая морфоген Б. Она будет расположена на строго определенном расстоянии от одного из полюсов зародыша, потому что в нашем примере морфоген А служит для изначальной разметки яйцеклетки. По концентрации морфогена А клетка может определить, на каком расстоянии от полюсов зародыша она находится.
Почему клетка, выделяющая морфоген Б, будет только одна? Потому что в инструкции есть такое условие: «Если концентрация морфогена Б равна нулю». Как только первая клетка, в которой выполнятся поставленные условия, начнет выделять вещество Б, его концентрация перестанет быть равной нулю, и поэтому другие клетки не начнут его выделять.
Что будет, если мы уберем из программы инструкцию «Если концентрация морфогена Б равна нулю»? Теперь программа поведения будет выглядеть так:
«Если ты снаружи, и если концентрация морфогена А такая-то, и если с момента начала развития пройдено десять делений, то начни выделять морфоген Б».
В этом случае морфоген Б начнет выделяться не одной клеткой, а целой полоской клеток, опоясывающей зародыш на определенном расстоянии от полюсов. Ширина этого пояса и его положение (ближе или дальше от того полюса, где концентрация А максимальна) будут зависеть от того, какие концентрации указаны в инструкции «Если концентрация морфогена А такая-то».
Чтобы сделать рассказ более наглядным, наш искушенный в программировании сын Михаил написал по нашей просьбе компьютерную программу EvoDevo. Она позволяет имитировать самосборку сложных многоклеточных структур из множества делящихся клеток. Программа получилась замечательная. Она дает нам одну начальную клетку — зиготу, а мы в специальном окошке пишем правила ее поведения . Например, такие, как в приведенном выше примере.
Мы можем задать любой набор правил поведения клетки. Следовать этим правилам будут все клетки эмбриона. Но мы не можем задать правило для всего эмбриона сразу. Мы не можем сказать эмбриону: «Втяни брюшко и прими форму шляпы». Мы можем задать только правила для клетки, например: «При таких-то условиях ползи туда, где возрастает концентрация такого-то вещества». И этот набор правил един для всех клеток. Правила — это генотип нашего модельного организма.
Написав генотип, мы нажимаем кнопку «старт» и смотрим, как из зиготы что-нибудь развивается. Например, описанный выше набор правил, который должен приводить к развитию зародыша с «пояском», породил вот такой симпатичный модельный эмбрион:
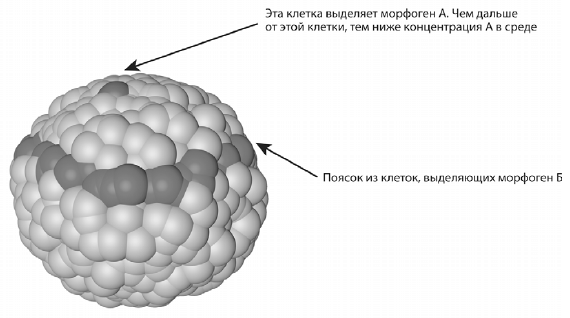
Поясок, правда, вышел неровный. Сделать его более аккуратным оказалось неожиданно трудно, хотя причина проста. Во всяком онтогенезе обязательно присутствует элемент стохастики (случайности). Полностью устранить случайность не удается ни естественному отбору, ни опытным пользователям программы EvoDevo, которая в этом отношении довольно точно воспроизводит реальность. Подробнее о стохастике в онтогенезе мы поговорим чуть ниже.
Как бы то ни было, теперь наш зародыш размечен гораздо сложнее и интереснее, чем в начале. У него есть передняя часть, в которой много А, а концентрация Б растет спереди назад (по направлению к пояску), сам центральный поясок, где максимальна концентрация Б, и задняя часть, где мало А и где концентрация Б убывает от пояска назад. Зародыш подразделился на четко отграниченные друг от друга части, в которых клетки находятся в разных условиях и поэтому могут выполнять разные подпрограммы своей исходной общей программы.
Мы подразделили зародыш на передний, средний и задний отделы. Они могут стать, например, головой, туловищем и хвостом. Но хотелось бы еще понять, где у него будет спина, а где брюшко. Как это сделать? Очень просто, мы уже это проделывали. Нужна инструкция, приводящая к появлению одной клетки или группы клеток, выделяющих какое-нибудь вещество (например, В) на любом «боку» зародыша, где-то посередине между головой и хвостом. И пусть это вещество В запустит программу развития спины там, где его много, и программу формирования брюха там, где его мало. Когда зародыш подробно размечен, каждая группа клеток может точно определить, где она находится, и активизировать заготовленную на этот случай подпрограмму (набор правил поведения).
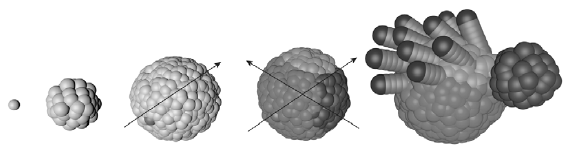 Пример развития двусторонне-симметричного организма в программеEvoDevo (из неполяризованной зиготы). Сначала в произвольном месте зародыша возникает одна клетка- организатор, выделяющая морфоген А. Градиентом концентрации этого морфогена(стрелка, направленная вправо и вверх) будет задаваться спинно-брюшная полярность (где его много, будет брюхо, где мало — спина). Затем на определенном расстоянии от первого организатора возникает второй — клетка, выделяющая морфоген Б. Градиент концентрации Б(стрелка, направленная влево и вверх) будет определять передне-заднюю полярность. После этого клетки зародыша, исходно одинаковые, подразделяются на три типа. Клетки, в которых концентрация Б максимальна, становятся «головными»(темными) . Все прочие клетки становятся либо «брюшными» (если концентрация А высока), либо «спинными» (если она низка). Из эмбриона, размеченного таким образом, можно вырастить разнообразные двусторонне-симметричные существа, добавляя в программу правила поведения, распространяющиеся только на клетки определенного типа. Такие правила должны начинаться с инструкции: «Если ты клетка такого-то типа, то…».
Пример развития двусторонне-симметричного организма в программеEvoDevo (из неполяризованной зиготы). Сначала в произвольном месте зародыша возникает одна клетка- организатор, выделяющая морфоген А. Градиентом концентрации этого морфогена(стрелка, направленная вправо и вверх) будет задаваться спинно-брюшная полярность (где его много, будет брюхо, где мало — спина). Затем на определенном расстоянии от первого организатора возникает второй — клетка, выделяющая морфоген Б. Градиент концентрации Б(стрелка, направленная влево и вверх) будет определять передне-заднюю полярность. После этого клетки зародыша, исходно одинаковые, подразделяются на три типа. Клетки, в которых концентрация Б максимальна, становятся «головными»(темными) . Все прочие клетки становятся либо «брюшными» (если концентрация А высока), либо «спинными» (если она низка). Из эмбриона, размеченного таким образом, можно вырастить разнообразные двусторонне-симметричные существа, добавляя в программу правила поведения, распространяющиеся только на клетки определенного типа. Такие правила должны начинаться с инструкции: «Если ты клетка такого-то типа, то…».
В ходе развития настоящего зародыша действительно то тут то там появляются — сами собой! — особые управляющие центры, так называемые организаторы. Никакой мистики, никакого разумного вмешательства. Организаторы — это просто группы клеток, выделяющие тот или иной морфоген, который служит для других клеток сигналом и влияет на их поведение. При этом все клетки по-прежнему ведут себя в строгом соответствии с изначальной генетической программой, которая у всех одна и та же. Управляющие центры возникают сами, путем самоорганизации, никто их нарочно туда не вставляет. Никакого «единого централизованного руководства» для этого не требуется. И его нет.
В развитии настоящих животных все сложнее, чем в придуманном нами примере, но, как ни странно, ненамного.
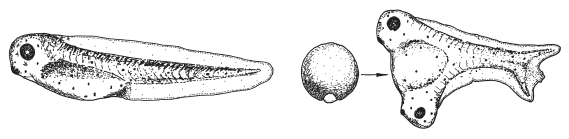
—————Организатор ШпеманаСтановление современной эволюционной биологии развития началось с поразительного результата, полученного в 1920-е годы эмбриологом Гансом Шпеманом (1869–1941) и его ученицей Хильдой Мангольд (1898–1924), трагически погибшей вскоре после этого эксперимента. Мангольд вырезала из зародыша тритона небольшой кусочек и пересадила его в другой зародыш, находящийся на той же стадии развития. В результате из второго зародыша развились два сросшихся тритона — сиамские близнецы. Как выяснилось, для получения такого результата необходимо брать кусочек для пересадки из определенного места первого зародыша — из спинной губы бластопора .Самое интересное, что «лишний» тритон развивается не из пересаженных клеток, а из клеток зародыша-реципиента (того, кому пересадили чужую ткань). В этом можно убедиться, если использовать в роли донора и реципиента зародыши тритонов, различающихся по окраске. Дополнительный тритон в этом случае будет окрашен так же, как и основной. Все гены у него будут такие же, как у реципиента: два тритона будут настоящими идентичными близнецами.Из этого следует, что спинная губа бластопора — не какая-то там «квинтэссенция» зародыша, содержащая в себе в концентрированном виде потенциал развития. Нет, спинная губа — это организатор, т. е. группа клеток, выделяющая вещества-морфогены, которые заставляют близлежащие клетки реципиента организоватьсяв тритона. Дальнейшие опыты подтвердили это: например, оказалось, что пересаживаемые клетки могут быть даже мертвыми. Главное, чтобы клетки реципиента подверглись действию содержащихся в них веществ.
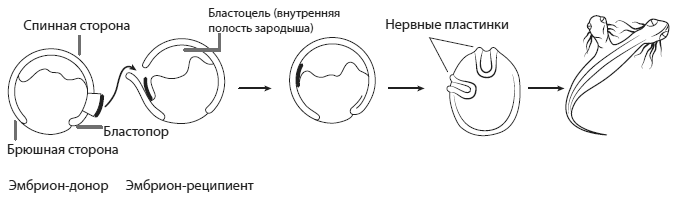 Эксперимент Шпемана — Мангольд. Пересадка фрагмента спинной губы бластопора стимулирует развитие «лишней» нервной пластинки рядом с пересаженным кусочком ткани, что, в свою очередь, индуцирует развитие «лишнего» организма.
Эксперимент Шпемана — Мангольд. Пересадка фрагмента спинной губы бластопора стимулирует развитие «лишней» нервной пластинки рядом с пересаженным кусочком ткани, что, в свою очередь, индуцирует развитие «лишнего» организма.
Очень долго (около полувека) эмбриологи тщетно пытались разгадать тайну «шпемановского организатора». В конце концов это удалось сделать. В предельно упрощенном виде принцип его действия сводится к выделению белка-морфогена, получившего название хордин (chordin). Этот белок присоединяется к другому морфогену, белку BMP4, производимому клетками будущей брюшной стороны зародыша. Тем самым хордин инактивирует BMP4 и не дает ему присоединиться к рецепторам соседних клеток эктодермы (наружного слоя зародыша). Там, где хордина мало (т. е. вдали от организатора), BMP4 соединяется с этими рецепторами, что заставляет клетки эктодермы превратиться в клетки кожи. Там, где хордина много (рядом с организатором), BMP4 не может присоединиться к рецепторам эктодермальных клеток. Поэтому они не становятся клетками кожи. Вместо этого они начинают развиваться по другой программе и становятся клетками нервной пластинки — зачатка центральной нервной системы. Нервная пластинка, в свою очередь, индуцирует развитие всех остальных тканей и органов будущего тритона.
 Результат аналогичного эксперимента с зародышем шпорцевой лягушки.
Результат аналогичного эксперимента с зародышем шпорцевой лягушки.
Шпемановский организатор играет ключевую роль в развитии позвоночных животных, и на самом деле его работа куда сложнее, чем здесь описано. Помимо ключевого взаимодействия «хордин инактивирует BMP4» спинно-брюшную полярность зародыша организует ряд дополнительных морфогенов. Их взаимодействия образуют сложную генно-регуляторную сеть с многочисленными обратными связями. Это необходимо для придания «программе развития» стабильности и помехоустойчивости. Мы еще вернемся к этой теме.—————
Как совершенно правильно (и очень внятно) объясняет в своих книгах Ричард Докинз, в геноме зиготы нет ничего похожего на чертеж организма. Между элементами чертежа и частями сработанного по нему изделия, как правило, существует соответствие «один к одному». Между частями генома и организма такого соответствия нет. Чуть точнее будет сравнение с рецептом (или алгоритмом), на основе которого повар готовит торт. Между частями пирога и отдельными элементами рецепта (словами, предложениями) нет взаимно однозначного соответствия. По готовому изделию можно восстановить его чертеж близко к оригиналу. Восстановить по «фенотипу» пирога его рецепт принципиально труднее, а восстановить его дословно в общем случае невозможно. Но метафора «рецепта» тоже несовершенна. Рецепт содержит команды на уровне целого изделия («придать тесту такую-то форму», «тщательно перемешать»). В геноме зиготы нет инструкций, выполняемых на уровне целого эмбриона. Геном содержит лишь набор локальных правил поведения, выполняемых на уровне отдельных клеток. Геном можно называть рецептом или алгоритмом только с этой оговоркой. В программировании такие алгоритмы известны. Их называют алгоритмами самосборки.
Если бы развитие шло не путем самоорганизации на основе программы, а по чертежу, нам было бы труднее эволюционировать. Лет 100 назад, когда мы еще не знали законов развития эмбриона, многое в эволюции казалось непонятным. Например, выдающийся биолог Лев Семенович Берг (1876–1950), автор теории номогенеза , сомневался, что за счет случайных мутаций могут удлиниться все четыре ноги одновременно — ведь для этого нужно, чтобы мутации одновременно и одинаково изменили длину сразу четырех органов! Иначе животное получится хромым. Действительно, если бы в геноме был записан чертеж организма, то потребовалось бы внести в этот чертеж четыре поправки, чтобы увеличить длину четырех ног. Но, поскольку развитие идет по программе, достаточно внести всего одно изменение, чтобы длина всех четырех конечностей изменилась согласованно. Разумеется, изменение-мутация должно быть внесено не куда попало, а именно в подпрограмму, контролирующую рост зачатков конечностей.
Скорее всего, закодировать полный чертеж животного намного сложнее, чем программу самосборки. Эта программа, как ни странно, сама по себе проще, чем получающийся в результате организм.
—————Пример самоорганизации: бактерии с полосатыми колониямиКак мы уже говорили, процессы самоорганизации, ведущие к самопроизвольному «рождению сложности», относятся к числу наиболее трудных для понимания. Поэтому очень полезны исследования, помогающие нам лучше представить себе механику самоорганизации.В 2011 году международная группа генетиков сообщила о создании генетически модифицированных бактерий E. coli, подвижность которых находится в обратной зависимости от плотности популяции ( Liu et al., 2011). В геном бактерий добавили несколько генов, составляющих два функциональных модуля: модуль определения плотности и модуль контроля подвижности. В роли первого выступает система « чувства кворума», заимствованная у светящейся бактерии Vibrio fischeri(с которой мы познакомились в главе 5). Бактерии выделяют сигнальное вещество AHL (ацил-гомосерин-лактон). Когда концентрация AHL в среде достигает порогового значения (это происходит при высокой плотности популяции), в бактериальных клетках активируется регуляторный белок LuxR.Модуль контроля подвижности эксплуатирует именно этот белок LuxR. Он работает следующим образом. У E. coliимеется ген cheZ, необходимый для нормального движения клеток. Бактерии без этого гена кувыркаются на месте и не могут передвигаться направленно. У генно-модифицированных бактерий белок LuxR, индуцированный веществом AHL, подавляет работу гена cheZ, и в результате бактерия кружится на месте.Когда модифицированных микробов посеяли на агаре, они стали образовывать колонии в виде правильных концентрических кругов. Светлые полосы соответствуют областям с высокой плотностью микробов. Полосы образуются последовательно, с постоянной скоростью, примерно по одной паре полос (светлая + темная) каждые четыре часа. Раз сформировавшись, полоса остается на своем месте. Обычные бактерии E. coli, как и контрольные микробы со встроенным модулем определения плотности, но с неизмененной системой регуляции cheZ, образуют равномерные круглые колонии без видимой структуры.
 Рост колонии генетически модифицированных бактерийE. coli . ИзLiu et al., 2011 .
Рост колонии генетически модифицированных бактерийE. coli . ИзLiu et al., 2011 .
Чтобы понять, почему модифицированные бактерии образуют полоски, авторам пришлось поставить ряд дополнительных экспериментов. В итоге выяснилось следующее. На первом этапе роста колонии плотность клеток низкая, и поэтому концентрация AHL не достигает порогового уровня, при котором клетки теряют способность к направленному движению. Примерно через 5 ч после начала роста в центре колонии достигается пороговая концентрация AHL, и бактерии в этой зоне теряют подвижность. Теперь они не могут покинуть эту область. В «зоне неподвижности» рост плотности клеток ускоряется. На границе этой зоны — там, где концентрация AHL по-прежнему ниже пороговой, — образуется область пониженной плотности бактерий. Дело в том, что бактерии, находящиеся возле границы, еще не потеряли подвижность, поэтому они свободно мигрируют во все стороны, в том числе и к центру колонии. В норме это компенсировалось бы обратной миграцией. Но клетки, попавшие в «зону неподвижности», сами становятся неподвижными и вернуться уже не могут. Это и приводит к формированию темной полосы с пониженной плотностью бактерий.Тем временем бактерии на «переднем фронте» растущей колонии продолжают спокойно размножаться и заселять новые территории. То, что происходит в центре колонии, не влияет на них, потому что концентрация AHL вокруг них пока остается низкой. Но через некоторое время снаружи от темной полосы (на строго определенном расстоянии от нее) возникает вторая зона с концентрацией AHL выше пороговой, и весь процесс повторяется: формируется новая светлая и новая темная полоса.
 Изысканные узоры, которые можно «нарисовать» модифицированными бактериями, если поселить их в двух, четырех или шести точках на питательной среде.Данное исследование может заинтересовать не только художников-авангардистов, которым пора задуматься о создании «саморисующихся» картин из генно-модифицированных бактерий . Результат может оказаться важным для понимания механизмов онтогенеза. Ведь в ходе развития многоклеточных различные повторяющиеся структуры формируются сплошь и рядом.
Изысканные узоры, которые можно «нарисовать» модифицированными бактериями, если поселить их в двух, четырех или шести точках на питательной среде.Данное исследование может заинтересовать не только художников-авангардистов, которым пора задуматься о создании «саморисующихся» картин из генно-модифицированных бактерий . Результат может оказаться важным для понимания механизмов онтогенеза. Ведь в ходе развития многоклеточных различные повторяющиеся структуры формируются сплошь и рядом.
При помощи математического моделирования авторы определили, что на частоту полос должны влиять такие факторы, как скорость распространения AHL в среде и подвижность бактерий. Полос будет меньше или они вообще не образуются, если AHL начнет распространяться в среде слишком свободно или если бактерии будут двигаться слишком вяло. Второе предсказание удалось проверить экспериментально. Для этого бактерий модифицировали еще раз, добавив в их геном дополнительный регулятор активности гена CheZ. Теперь активность этого гена зависела не только от плотности популяции бактерий, но и от концентрации ангидротетрациклина — вещества, которое экспериментаторы могли произвольно добавлять в среду. Модельное предсказание подтвердилось. Снижение подвижности бактерий (при концентрации AHL ниже пороговой) привело сначала к тому, что бактерии вместо бесконечного числа полос стали образовывать только 3–4 полосы, а затем росли равномерно. Дальнейшее уменьшение подвижности привело к полному исчезновению полос: теперь модифицированные бактерии росли как обычные E. coli.—————
Назад: Глава 8 Эволюция алгоритмов
Дальше: Неустранимая случайность

