Книга: Эволюция. Классические идеи в свете новых открытий
Назад: «Ничего особенно не трудно, если разделить работу на части»
Дальше: Глава 8 Эволюция алгоритмов
Сначала чутье, потом философия
Мозг млекопитающих (и особенно человека) нередко сравнивают со сверхсложным компьютером, способным выполнять сразу множество задач. Создать подобный компьютер и довести его до такого совершенства, чтобы он работал, не «повисая» каждую минуту, и верно служил своему обладателю всю его жизнь… ну как такое сделать! Однако природа все же справилась с этим, постепенно наращивая мощности и координируя когнитивные задачи. Начав с нервной трубки первых хордовых, естественный отбор постепенно добрался до млекопитающих с крупным мозгом и развитым неокортексом (новой корой).
Мозг млекопитающих радикально отличается от мозга рептилий не только своим размером, но и строением. В частности, у млекопитающих резко увеличились обонятельные луковицы и отделы коры, связанные с обонянием, а также мозжечок.
Особенно нас занимает процесс формирования неокортекса, позволяющего нам, читателям, разбирать и воспринимать буковки, складывать их в слова, анализировать информацию, а затем, запоминая ее, участвовать в дальнейших дебатах. Так как же мы, млекопитающие, получили этот изумительно сложный орган — неокортекс, кору больших полушарий?
Непосредственных предков первых млекопитающих относят к группе маммалиаформ, которые, в свою очередь, берут начало от цинодонтов. Палеонтологи детально реконструировали этапы эволюционных преобразований зубов и скелета цинодонтов по мере их «маммализации» — постепенного эволюционного движения в сторону млекопитающих. Об эволюции мозга известно значительно меньше. Между тем очевидно, что именно развитие мозга во многом предопределило эволюционный успех млекопитающих.
Изучение мозга маммалиаформ и первых млекопитающих затруднялось, во-первых, тем, что редко удавалось найти хорошо сохранившийся череп, во-вторых — тем, что для изучения эндокрана (слепка мозговой полости, по которому можно судить о размере и форме мозга) череп, как правило, нужно было разрушить.
Американским палеонтологам удалось в некоторой мере восполнить этот досадный пробел ( Rowe et al., 2011). При помощи компьютерной рентгеновской томографии, не разрушая драгоценных черепов, они получили детальные трехмерные изображения эндокранов двух маммалиаформ, живших в начале ранней юры (200–190 млн лет назад) на территории нынешнего Китая.
Изученные маммалиаформы Morganucodon oehleri(уже упоминавшееся животное с готовыми косточками среднего уха) и Hadrocodium wui— ближайшие родственники первых млекопитающих. По строению скелета они представляют собой классические переходные формы между «еще рептилиями» и «уже млекопитающими». При этом Morganucodonстоит ближе к примитивным цинодонтам, а Hadrocodiumнастолько близко подошел к млекопитающим, насколько это возможно, оставаясь формально за пределами группы. Исследование показало, что по строению мозга эти животные тоже занимают промежуточное положение между типичными цинодонтами и их потомками — млекопитающими.
Ранее были изучены эндокраны ранних цинодонтов Thrinaxodonи Diademodon. Выяснилось, что мозг у них был еще вполне рептильный по своему размеру и строению.
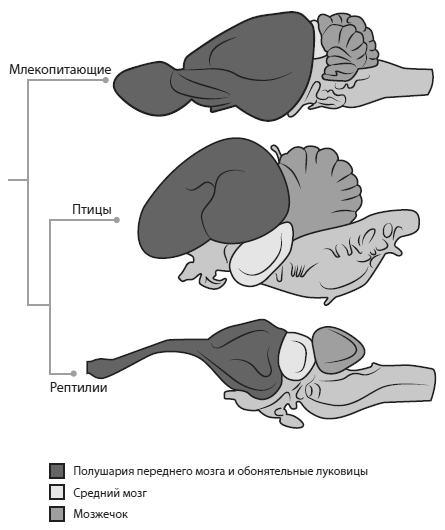 Соотношение отделов мозга у млекопитающих, птиц и рептилий.
Соотношение отделов мозга у млекопитающих, птиц и рептилий.
Относительный размер мозга у позвоночных оценивается при помощи «коэффициента энцефализации» (EQ), который вычисляют по эмпирически выведенной формуле EQ = EV/(0,055[Wt] 0,74), где EV — объем мозговой полости в миллилитрах, Wt — масса тела в граммах.
У базальных цинодонтов Thrinaxodonи DiademodonEQ варьирует от 0,16 до 0,23. Для сравнения, у крысы EQ = 0,53. Обонятельные луковицы у них были небольшие, а в носу не было окостеневших носовых раковин, что свидетельствует о слабом развитии обонятельного эпителия. Передний мозг был маленький и узкий, не подразделенный на отделы, без признаков наличия неокортекса. Средний мозг и эпифиз (теменной глаз) не были закрыты сверху полушариями переднего мозга. Мозжечок был шире переднего мозга, спинной мозг тонкий. Эти и другие «рептильные» особенности мозга и черепа цинодонтов свидетельствуют о том, что у них по сравнению с млекопитающими было слабое обоняние и не слишком совершенный слух, осязание и координация движений.
 Morganucodon.
Morganucodon.
Мозг Morganucodon, как выяснилось, был больше похож на мозг млекопитающего. По объему он в полтора раза превосходит мозг базальных цинодонтов (EQ = 0,32). Сильнее всего увеличилась обонятельная луковица и обонятельная кора. Это со всей очевидностью указывает на более развитое обоняние. Полушария переднего мозга стали выпуклыми благодаря развитию неокортекса. Они закрывают сверху средний мозг и эпифиз, как у млекопитающих. Передний мозг морганукодона шире мозжечка, хотя мозжечок тоже заметно вырос по сравнению с базальными цинодонтами. Увеличение мозжечка говорит об улучшенной координации движений. На это же указывает и более толстый, чем у базальных цинодонтов, спинной мозг.
 Базальный цинодонтThrinaxodon (Южная Африка, Антарктика, ранний триас, 248–245 млн лет назад).
Базальный цинодонтThrinaxodon (Южная Африка, Антарктика, ранний триас, 248–245 млн лет назад).
Развитие неокортекса у древних млекопитающих было связано в первую очередь с совершенствованием соматосенсорных функций. Значительная часть неокортекса у примитивных млекопитающих, таких как опоссум, — это так называемая соматосенсорная кора, которая отвечает за сбор и анализ сигналов, приходящих от многочисленных механорецепторов, рассеянных по всему телу. Особенно много таких рецепторов приурочено к волосяным фолликулам.
По мнению многих палеонтологов, волосы выполняли сначала тактильную (осязательную) функцию, а для терморегуляции стали использоваться позже, когда у предков млекопитающих начала развиваться теплокровность. У Morganucodonи Hadrocodiumдостоверных остатков волосяного покрова пока не обнаружено, но их близкий родственник — похожий на бобра маммалиаформ Castorocauda(в переводе с латыни — «с хвостом как у бобра») — был покрыт густым мехом, состоявшим из осевых волос и подшерстка. Это позволяет предположить, что Morganucodonи Hadrocodiumтоже были покрыты шерстью. Вероятно, появление неокортекса у маммалиаформ было связано в том числе и с развитием волосяного покрова и осязания.
Таким образом, мозг примитивного Morganucodonиллюстрирует первый этап прогрессивной эволюции мозга в ходе становления млекопитающих. На этом этапе увеличение мозга было обусловлено развитием обоняния, осязания и координации движений.
Hadrocodium, более продвинутый представитель маммалиаформ и ближайший родственник «настоящих» млекопитающих, иллюстрирует второй этап развития мозга. Коэффициент энцефализации у Hadrocodiumравен 0,5, т. е. мозг увеличился еще в полтора раза по сравнению с Morganucodonи достиг размеров, характерных для некоторых настоящих млекопитающих. Мозг вырос в основном за счет обонятельных луковиц и обонятельной коры. Таким образом, второй этап прогрессивной эволюции мозга в ходе маммализации тоже был связан с развитием обоняния.
Третий этап соответствует переходу от высших маммалиаформ, таких как Hadrocodium, к настоящим млекопитающим. На этом этапе обоняние становится еще более тонким, о чем свидетельствуют изменения решетчатой кости: на ней образуются носовые раковины, поддерживающие разросшийся обонятельный эпителий.
Эти данные показывают, что потребность в тонком чутье была, по-видимому, главным стимулом развития мозга в ходе становления млекопитающих. У млекопитающих, как известно, обоняние развито намного лучше, чем у всех остальных наземных позвоночных. Скорее всего, изначально это было связано с приспособлением к ночному образу жизни, когда на зрение полагаться бессмысленно (мы говорили об этом в главе 5). В такой ситуации было выгодно совершенствовать уже имеющуюся древнюю систему обоняния: разнообразить обонятельные рецепторы и усиливать мозговые мощности для их анализа. Что ж, пусть это было обоняние, зато в качестве побочного продукта млекопитающие получили кору с массой нейронов и в конце концов стали использовать ее для осмысления произошедших перемен.
Назад: «Ничего особенно не трудно, если разделить работу на части»
Дальше: Глава 8 Эволюция алгоритмов

