Книга: Эволюция. Классические идеи в свете новых открытий
Назад: Дупликация генов
Дальше: Откуда взялся новый фермент?
Секретный код позвоночных выдал ланцетник
Ланцетник ( Branchiostoma) — хрестоматийный пример живого ископаемого, т. е. мало изменившегося потомка очень древнего предка. Ланцетника называют живой схемой хордового животного. Это маленькое червеобразное существо большую часть времени проводит, зарывшись в грунт и отфильтровывая пищевые частицы из морской воды. Вода входит в рот, а выходит через жаберные щели, соединяющие полость глотки с окружающей средой. Ланцетник удивительно похож на ранние стадии эмбрионального развития позвоночных. Это было замечено еще в XIX веке эмбриологом А. О. Ковалевским (1840–1901). Как и у зародышей позвоночных, у ланцетника есть спинная струна — хорда, спинная нервная трубка, жаберные дуги, сердце на брюшной стороне тела и другие характерные признаки, доказывающие его родство с позвоночными.
Тип хордовых делится на три подтипа: головохордовые (единственным представителем которых является ланцетник), оболочники (асцидии, сальпы, аппендикулярии) и позвоночные. Родственные связи подтипов до недавних пор оставались не совсем ясными. Одни эксперты считали, что ланцетник ближе к позвоночным, чем оболочники, другие утверждали, что ближайшими родственниками позвоночных являются оболочники. Впрочем, по поводу того, как примерно выглядел общий предок хордовых, особых разногласий нет уже со времен Ковалевского. Это было червеобразное животное с хордой, спинной нервной трубкой и жаберными щелями, сходное с ланцетником, личинками оболочников и пескоройками — личинками миног. Выводы сравнительной анатомии и эмбриологии подкрепляются палеонтологическими данными: отпечатки примитивных хордовых животных, напоминающих ланцетника, обнаружены в отложениях кембрийского возраста. Как раз в это время и должны были жить формы, близкие к общему предку трех современных подтипов.
В последнее время для выяснения спорных вопросов ранней эволюции хордовых активно привлекаются молекулярногенетические данные. Сначала работали с отдельными генами, потом в распоряжении ученых оказались полные тексты геномов различных позвоночных, а затем и оболочников. Эволюционные деревья постепенно становились все более точными. Однако для окончательного решения ряда вопросов не хватало генома ланцетника. Наконец в 2008 году он был прочтен ( Putnam et al., 2008).
Размер генома ланцетника — 520 млн пар нуклеотидов (в шесть раз меньше, чем у человека); в нем содержится 21 900 белок-кодирующих генов (у человека — примерно столько же); мобильные генетические элементы составляют 30 % генома (у человека — свыше 50 %).
Ланцетники, как выяснилось, генетически очень разнообразны. Парные хромосомы, которые изученный ланцетник получил от отца и матери, различаются по своим нуклеотидным последовательностям на 3,7 % (если учитывать только нуклеотидные замены; вставки и выпадения нуклеотидов добавляют еще 6,8 % различий). Это значит, что родители этого ланцетника генетически отличались друг от друга втрое сильнее, чем человек от шимпанзе, и в десятки раз сильнее, чем отличаются друг от друга представители человеческих рас и народов. Высокий полиморфизм, скорее всего, объясняется тем, что численность популяции, к которой принадлежал исследованный ланцетник, оставалась очень высокой в течение долгого времени. Ланцетники действительно являются массовыми животными. В Китае, например, их гребут лопатами (в буквальном смысле), промывают от песка и варят суп.
Прочтенный геном ланцетника сыграл роль последнего недостающего фрагмента в сложной головоломке родственных отношений вторичноротых животных (к вторичноротым относят хордовых, полухордовых и иглокожих). До его прочтения эволюционное дерево вторичноротых сохраняло долю неопределенности. Но теперь, похоже, для сомнений больше не осталось места. Авторы использовали для построения дерева 1090 генов, общих для всех животных, — больше просто некуда. И дерево получилось весьма устойчивым (каждая точка ветвления имеет очень высокую «статистическую поддержку»). Поэтому можно уверенно сказать, что дерево, показанное на рисунке, уже не будет в дальнейшем меняться. Это окончательное эволюционное дерево вторичноротых, показывающее (даже страшно вымолвить!) истинный ход их эволюции.
Вторичноротые сначала разделились на хордовых и предков иглокожих и полухордовых. От эволюционного ствола хордовых первыми отделились предки ланцетника и только потом — предки оболочников. Таким образом, оболочники все-таки оказались более близкими родственниками позвоночных, чем ланцетник. Зато ланцетник подтвердил свое право на почетное звание самого примитивного хордового (точнее, как сейчас принято говорить, раньше всех ответвившегося).
Еще в 1970 году американский биолог Сусуму Оно, автор монографии «Эволюция путем дупликации генов» ( Ohno, 1970), предположил, что на заре эволюции хордовых их геном подвергся одной или двум дупликациям. Избыточность генома резко возросла, «лишние» гены получили возможность эволюционировать в новых направлениях, что открыло позвоночным путь к созданию сложных адаптаций.
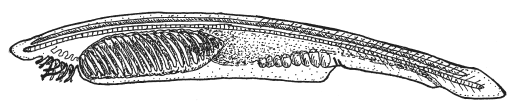 Ланцетник — «живая упрощенная схема хордового животного».
Ланцетник — «живая упрощенная схема хордового животного».
Гипотеза Оно сначала основывалась на скудных фактах, но постепенно появлялись все новые аргументы в ее пользу. В итоге она стала почти общепризнанной, но не хватало решающего доказательства. Ученые возлагали надежды на геномы оболочников, но тут их постигло разочарование. Оказалось, что молекулярная эволюция оболочников протекала исключительно бурно (это видно, в частности, по длине соответствующих веточек на рисунке). В результате геномы оболочников оказались очень сильно перемешаны. Гены оболочников так часто переходили с места на место, с одной хромосомы на другую, что от исходного порядка расположения генов в хромосомах мало что осталось. Между тем для решения вопроса о том, были ли в эволюции хордовых геномные дупликации, необходим анализ «групп сцепления» — длинных участков ДНК, содержащих сходные наборы генов у разных животных. Хромосомы в ходе эволюции рвутся, склеиваются и обмениваются участками, но отдельные кусочки, в том числе довольно длинные, могут сохраняться в относительной целости сотни миллионов лет. Сопоставляя геном ланцетника с геномами позвоночных, ученым удалось реконструировать 17 «исходных групп сцепления», которые соответствуют 17 хромосомам общего предка хордовых. Это означает, что у ланцетника нашлись такие группы генов, которые расположены сходным образом и у позвоночных. Это следы того исходного порядка, в котором гены располагались у последнего общего предка хордовых.
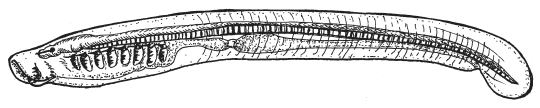 Пескоройка — личинка миноги — представляет собой, как и ланцетник, живую упрощенную схему типичного хордового. Она не сосет кровь у рыб, как многие взрослые миноги, а фильтрует воду через жаберные щели, подобно ланцетнику.
Пескоройка — личинка миноги — представляет собой, как и ланцетник, живую упрощенную схему типичного хордового. Она не сосет кровь у рыб, как многие взрослые миноги, а фильтрует воду через жаберные щели, подобно ланцетнику.
Как и следовало ожидать, каждая из 17 предковых хромосом превратилась в геноме его потомков (в том числе человека) в набор обрывков, рассеянных по разным хромосомам. Теперь мы подошли к самому главному. Оказалось, что каждый из 17 наборов в свою очередь подразделяется на четыре части. Например, набор, соответствующий предковой хромосоме № 8, в геноме человека состоит из девяти фрагментов ДНК. Они подразделяются на четыре группы (в трех группах по два фрагмента, в четвертой — три). Если сравнить между собой этих четыре группы фрагментов, то выясняется, что многие гены представлены в нескольких из них в виде паралогов(или паралогичных генов), по одному паралогу в группе. Паралогичные гены — это родственные гены, когда-то произошедшие от общего гена-предка в результате дупликации. Таким образом, каждая предковая хромосома превратилась в геноме позвоночных в четыре сходные между собой группы фрагментов ДНК. Сходство заключается в том, что если в одной группе есть какой-то ген, то в трех других с высокой вероятностью найдутся его паралоги. Это можно объяснить только тем, что на ранних этапах эволюции позвоночных действительно произошло учетверение генома — две последовательные дупликации. Гипотезу Оно, высказанную более 40 лет назад, наконец можно считать доказанной.
Дупликации произошли после того, как предки хордовых отделились от предков ланцетников и оболочников (их геномы не учетверены и не удвоены), и до того, как разошлись пути костных и хрящевых рыб. Скорее всего, первое удвоение произошло незадолго до, а второе сразу после разделения линий бесчелюстных (миноги, миксины) и челюстноротых (рыбы и наземные позвоночные). Эти линии разделились в раннем кембрии, примерно 530 млн лет назад.
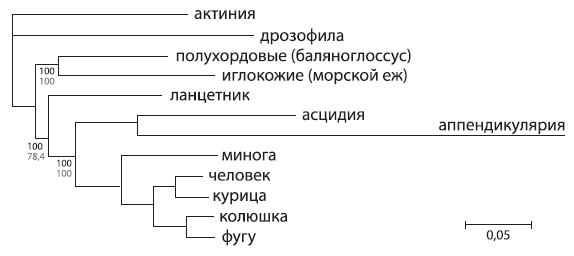 Эволюционное дерево вторичноротых, основанное на последовательностях 1090 белков. Актиния и дрозофила использованы в качестве «внешних групп», асцидия и аппендикулярия представляют оболочников. Длина ветвей отражает количество аминокислотных замен. ИзPutnam et al., 2008 .
Эволюционное дерево вторичноротых, основанное на последовательностях 1090 белков. Актиния и дрозофила использованы в качестве «внешних групп», асцидия и аппендикулярия представляют оболочников. Длина ветвей отражает количество аминокислотных замен. ИзPutnam et al., 2008 .
Удвоение целого генома — далеко не уникальный случай в истории жизни. Еще одна полногеномная дупликация произошла 350 млн лет назад у предков костистых рыб; 40 млн лет назад то же самое случилось в одной из групп лягушек. Обычно после этого большинство «лишних» генов быстро теряется, и лишь некоторым из них находится новое применение. До тех пор пока два гена-паралога не начнут выполнять в организме разные функции, естественный отбор не может воспрепятствовать мутационной поломке или утрате одного из них.
Ученые составили список из 8437 генов, которые были у общего предка хордовых и сохранились хотя бы у некоторых современных позвоночных. Все эти гены в свое время учетверились, но большая часть появившихся паралогов была утрачена. Сохранились паралоги у 20–25 % генов. Среди генов, дубликаты которых «пригодились» ранним позвоночным, резко повышена доля тех, которые участвуют в регуляции транскрипции (гены — регуляторы активности других генов), в работе нервной системы, в регуляции эмбрионального развития и в передаче разнообразных сигналов. Это явно неслучайный выбор. Он хорошо согласуется с идеей о том, что геномные дупликации у предков позвоночных стали предпосылкой для последующей эволюции по пути усложнения.
Но повторим — естественный отбор не умеет заглядывать в будущее. Он не мог заранее знать, что позвоночные собираются эволюционировать именно в сторону усложнения. Здесь уместно вспомнить о механизме «ухода от адаптивного конфликта», о котором мы говорили выше. Не потому ли сохранились дубликаты генов-регуляторов, что уже у древнейших позвоночных эти гены были многофункциональными и не могли толком оптимизироваться для выполнения какой-то одной формообразующей функции? По-видимому, полногеномные дупликации предоставили позвоночным уникальную возможность быстро снять многие из накопившихся адаптивных конфликтов и, как следствие, избавиться от множества запретов, стоявших на пути морфологических преобразований.
Назад: Дупликация генов
Дальше: Откуда взялся новый фермент?

