Книга: Эволюция. Классические идеи в свете новых открытий
Назад: Изменения белок-кодирующих генов
Дальше: Дупликация генов
Изменения регуляции генов
Не всякое новшество требует изменений белок-кодирующих участков ДНК. В основе многих эволюционных преобразований лежат мутации регуляторных некодирующих участков, от которых зависит, где, когда, при каких условиях и с какой интенсивностью будет работать близлежащий ген. Изучать такие эволюционные изменения технически намного труднее, но и на этом фронте биологи в последние годы добились важных успехов.
ЗАГАДКА УЗОРЧАТЫХ КРЫЛЬЕВ.
Плодовые мушки рода Drosophila— идеальный объект для изучения эволюции сложных признаков. В этой группе перемешаны не только узкие и широкие экологические специалисты, но и виды с разнообразными морфологическими признаками. Причем характер родственных связей между видами установлен вполне надежно (т. е. известно, кто от кого и когда произошел), и можно проследить, как в ряду потомков видоизменялся тот или иной сложный признак.
Например, у D. guttiferaна крыльях имеется замысловатый узор, которого нет у других дрозофил. Узор состоит из 16 черных пятен, расположенных в строго определенных местах на продольных жилках и в точках соединения продольных жилок с поперечными, и нескольких серых «теней» между жилками. У родственников D. guttiferaкрылья либо вовсе не окрашены, либо имеют гораздо более простой орнамент. Очевидно, сложная окраска крыльев D. guttifera— недавнее эволюционное приобретение.
Шон Кэрролл, американский генетик и автор превосходных научно-популярных книг, и его коллеги из Висконсинского университета решили выяснить, какие генетические изменения ответственны за появление у D. guttiferaзатейливого орнамента на крыльях. Ранее эта исследовательская группа установила, что независимое появление простых темных пятен на крыльях у разных видов дрозофил связано с изменениями регуляторных участков гена yellow, который управляет синтезом пигмента меланина и отвечает за пигментацию разных частей тела. Об этом исследовании рассказано в книге «Рождение сложности» ( Марков, 2010. С. 344).
Выяснилось, что на стадии куколки yellowработает как раз в тех участках крыла, где впоследствии появятся темные пятна. Следовательно, у этого вида, как и у других дрозофил, формирование пятен идет под управлением yellow. Однако у D. guttiferaраспределение областей экспрессии yellowпо крылу отличается особой сложностью.
Нужно было понять, что заставляет yellowработать в одних частях крыла и не работать в других. Зная, что узоры на крыльях дрозофил зависят от некодирующих регуляторных участков yellow, авторы принялись искать эти участки . Они отсеквенировали фрагмент генома D. guttiferaдлиной в 42 тыс. пар нуклеотидов, включающий ген yellowи его окрестности. Затем они подразделили некодирующие области на 28 перекрывающихся кусочков и стали изучать их свойства при помощи генно-инженерных экспериментов. Каждый кусочек по очереди присоединяли к гену зеленого флуоресцирующего белка (GFP) и вставляли эту конструкцию в геном D. guttifera, а затем смотрели, где будет вырабатываться GFP Таким непростым способом удалось выявить регуляторный участок длиной в 277 пар нуклеотидов, который заставляет присоединенный к нему ген включаться как раз в тех 16 точках развивающегося крыла, где у диких мух D. guttiferaвключается ген yellowи формируются пятна на жилках. Этот регуляторный участок находится на расстоянии около 5 тыс. пар нуклеотидов перед началом кодирующей части yellow. Таким образом, один-единственный регуляторный элемент контролирует формирование всех 16 пятен.
Тем же способом был выявлен еще один регуляторный участок, который отвечает за серые «тени» между жилками. Этот участок длиной в 414 пар нуклеотидов расположен в первом интроне гена yellow. Если присоединить его к гену светящегося белка и вставить в геном D. guttifera, у куколки начинают светиться те участки крыла, где должны быть «тени».
Следовательно, всего лишь два регуляторных элемента определяют все распределение областей экспрессии yellowпо крыловой пластинке (хотя в принципе каждое пятнышко могло бы определяться своим собственным регуляторным элементом). Первый из обнаруженных элементов авторы назвали vs( vein spot— «пятна на жилках»), второй — iv( intervein shade— «тени между жилками»). Эти элементы действуют как переключатели. Первый из них срабатывает там, где должны сформироваться пятна, второй — в местах образования будущих «теней».
Таким образом, «нижняя» часть регуляторного каскада была в общих чертах расшифрована. Распределение пятен по крылу определяется элементами vsи iv. Срабатывание переключателей vsи ivактивирует ген yellow. Ген yellowвключает синтез меланина. Синтез меланина приводит к формированию пятна на крыле.
Этого уже было бы достаточно для хорошей статьи. Но авторы не остановились на достигнутом и продвинулись еще на один шаг вверх по регуляторному каскаду. Им удалось выяснить, на какой сигнал реагирует переключатель vs— что заставляет его включаться в нужном месте и в нужное время.
Не имея никаких подсказок, подступиться к такой задаче непросто, ведь в принципе присоединяться к элементу vs, запуская экспрессию yellow, мог бы любой из многих сотен регуляторных белков (транскрипционных факторов), а на работу этого неизвестного белка наверняка влияют еще какие-то факторы. Авторы, однако, добыли подсказку, изучая отклонения от нормального хода развития в лабораторной популяции мух D. guttifera. Они заметили, что пятна жестко привязаны к определенным опорным точкам на крыловой пластинке. Такими опорными точками являются поперечные жилки и места их соединения с продольными жилками, окончания продольных жилок, а также колоколовидные сенсиллы — рецепторы, расположенные на продольных жилках. Если у мутантной мухи на крыле появляется новая сенсилла или новое место соединения жилок, это всегда приводит к формированию нового пятна. Если, наоборот, сенсилла теряется, то исчезает и соответствующее пятно.
Следовательно, сложный рисунок на крыльях связан с предшествующей «разметкой» крыловой пластины, которая определяет ее структуру. Это позволило сузить круг поиска. Генетическая разметка развивающегося эмбриона или куколки осуществляется небольшим набором генов — ключевых регуляторов развития. Возможно, какой-то из них взаимодействует — прямо или опосредованно — с переключателем vs.
Чтобы найти искомый ген, авторы пересадили элемент vs, присоединенный к гену GFP, в геном мухи D. melanogaster— классического лабораторного объекта, чья генетика развития изучена гораздо лучше, чем у D. guttifera. Оказалось, что у D. melanogasterэлемент vsсрабатывает у основания крыла и на поперечных жилках, но не работает вблизи колоколовидных сенсилл. Точно так же распределены у этого вида и области экспрессии гена wingless— одного из важнейших регуляторов индивидуального развития. На ранних этапах онтогенеза членистоногих winglessучаствует в глобальной разметке туловища, формировании конечностей, определяет границы сегментов. На поздних стадиях развития куколки winglessзанят менее грандиозными проектами, участвуя в том числе в оформлении некоторых деталей строения крыльев.
Пересаженный от D. guttiferaэлемент vsсработал в крыле куколки D. melanogasterровно в тех местах, где активен ген wingless. Это делает winglessхорошим кандидатом на роль «входного сигнала», активирующего vsи включающего yellow. Авторы проверили, как распределены области экспрессии winglessв крыле D. guttifera, и убедились, что все сходится: у вида с пятнистыми крыльями winglessна стадии куколки работает как раз в тех участках крыла, где позже формируются пятна.
Чтобы окончательно убедиться в том, что активность winglessявляется достаточным условием для появления пятен у D. guttifera, авторы провели еще одну серию генно-инженерных экспериментов. На этот раз в эмбрионы D. guttiferaвпрыскивали ген wingless, взятый у D. melanogasterи соединенный с регуляторным участком, заимствованным у дрожжей. Смысл заключался в том, чтобы получить трансгенных мух D. guttifera, у которых области экспрессии winglessв крыльях были бы расположены как-то иначе. В конце концов такие мухи были получены: у них возникла новая область экспрессии winglessвдоль одной из продольных жилок. В полном соответствии с теоретическими ожиданиями у этих мух появилась и новая темная полоса на крыльях, идущая вдоль этой жилки.
Авторы также проанализировали пигментацию крыльев, экспрессию winglessи наличие или отсутствие регуляторного элемента vsу других видов дрозофил. В итоге удалось восстановить ход эволюции узора на крыльях.
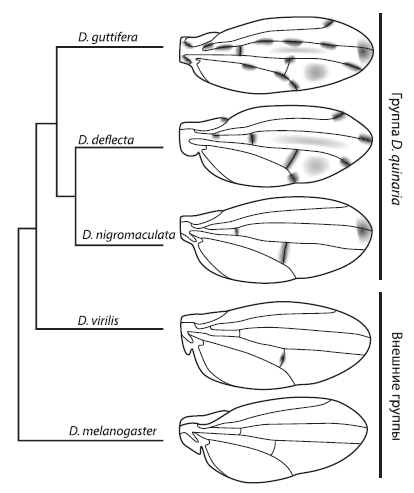 Слева — эволюционное дерево, показывающее родственные отношения между видами дрозофил с различными узорами на крыльях. Самый сложный орнамент — уD. guttifera . Справа — схема эволюции узора. ИзWerner et al., 2010 .
Слева — эволюционное дерево, показывающее родственные отношения между видами дрозофил с различными узорами на крыльях. Самый сложный орнамент — уD. guttifera . Справа — схема эволюции узора. ИзWerner et al., 2010 .
Исходно у дрозофил ген winglessэкспрессировался вдоль поперечных жилок, но не работал у концов продольных жилок и в колоколовидных сенсиллах. В гене yellowне было регуляторного элемента vs, способного реагировать на wingless, а крылья не имели орнамента. Это исходное состояние сохранилось у D. melanogaster. Затем в одной из групп дрозофил появился элемент vs, что привело к формированию связи между экспрессией winglessи пятнами на крыльях . Первые темные пятна поэтому появились там, где исходно экспрессировался wingless, — вдоль поперечных жилок.
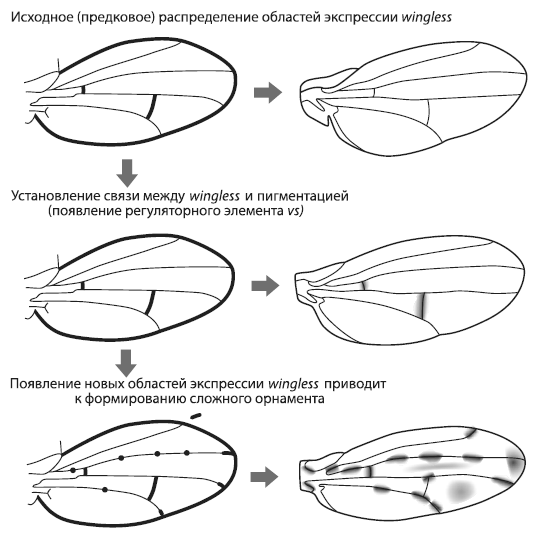
После того как пигментация стала зависеть от wingless, у эволюции появилась возможность создавать новые узоры, меняя экспрессию этого гена. Ген winglessимеет сложную и пока еще слабо изученную систему регуляции, но в целом его работа ориентируется на базовую разметку и ключевые «опорные точки» развивающегося организма. Эта привязанность winglessк опорным точкам сохранилась и в ходе дальнейших изменений его экспрессии в крыльях. У некоторых видов возникли новые области экспрессии winglessна концах продольных жилок, и там появились пятна. Только у одного вида — D. guttifera— орнамент крыльев усложнился еще сильнее за счет появления дополнительных областей экспрессии winglessв шести колоколовидных сенсиллах.
Получается, что замысловатый орнамент крыльев D. guttiferaразвился на основе существовавшего ранее плана строения крыла. Возможно, примерно таким же образом, отталкиваясь от опорных точек морфогенетической разметки крыла, сформировались узоры на крыльях и у других насекомых, включая бабочек. Но, чтобы это доказать, придется еще немало потрудиться.
Здесь мы видим, как изменение двух регуляторных участков одного гена (гена yellow) привело к формированию сложного признака. Изменение это было таково, что направило уже существующий регуляторный каскад по новому маршруту: старые персонажи в новой пьесе.
—————Как трехкамерное сердце стало четырехкамернымВыход позвоночных на сушу был связан с развитием легочного дыхания, что потребовало радикальной перестройки кровеносной системы. У дышащих жабрами рыб один круг кровообращения, а сердце, соответственно, двухкамерное (состоит из одного предсердия и одного желудочка). У наземных позвоночных — трех- или четырехкамерное сердце и два круга кровообращения. Один из них (малый) прогоняет кровь через легкие, где она насыщается кислородом. Затем кровь возвращается к сердцу и попадает в левое предсердие. Большой круг направляет обогащенную кислородом (артериальную) кровь ко всем прочим органам, где она отдает кислород и по венам возвращается к сердцу, попадая в правое предсердие.У животных с трехкамерным сердцем кровь из обоих предсердий попадает в единый желудочек, откуда она затем направляется и к легким, и ко всем прочим органам. При этом артериальная кровь смешивается с венозной. У животных с четырехкамерным сердцем в ходе развития изначально единый желудочек подразделяется перегородкой на левую и правую половины. В результате два круга кровообращения оказываются полностью разделены: бедная кислородом кровь попадает из правого предсердия в правый желудочек и идет оттуда к легким, насыщенная кислородом — из левого предсердия только в левый желудочек и идет оттуда ко всем прочим органам.Формирование четырехкамерного сердца было необходимой предпосылкой развития теплокровности у млекопитающих и птиц. Ткани теплокровных потребляют очень много кислорода, поэтому им необходима «чистая» артериальная кровь, максимально насыщенная кислородом. А смешанной артериально-венозной кровью могут довольствоваться холоднокровные позвоночные с трехкамерным сердцем. Трехкамерное сердце характерно для амфибий и большинства рептилий, хотя у последних намечается частичное разделение желудочка на две части (развивается неполная внутри-желудочковая перегородка). Настоящее четырехкамерное сердце развилось независимо в трех эволюционных линиях: у крокодилов, птиц и млекопитающих. Это яркий пример параллельной эволюции.Биологам из США, Канады и Японии удалось частично расшифровать молекулярно-генетические основы этого важнейшего эволюционного события ( Koshiba-Takeuchi et al., 2009). Ключевую роль в нем сыграли изменения в работе гена Tbx5. Этот ген, кодирующий регуляторный белок, по-разному экспрессируется в развивающемся сердце у амфибий (шпорцевой лягушки Xenopus) и теплокровных (курицы и мыши). У первых он равномерно экспрессируется по всему будущему желудочку, у вторых его экспрессия максимальна в левой части зачатка (в будущем левом желудочке) и минимальна справа. А как обстоит дело у рептилий?Выяснилось, что у рептилий — ящерицы и черепахи — на ранних эмбриональных стадиях ген Tbx5экспрессируется так же, как у лягушки, т. е. равномерно по всему будущему желудочку. У ящерицы все так и остается до конца развития. Как и у лягушки, у ящерицы не формируется ничего похожего на перегородку (хотя бы частичную) между желудочками.Что касается черепахи, то у нее на поздних стадиях формируется градиент экспрессии — такой же, как у цыпленка, только выраженный слабее. Иными словами, в правой части желудочка активность гена постепенно снижается, а в левой остается высокой. Таким образом, по характеру экспрессии Tbx5черепаха занимает промежуточное положение между ящерицей и курицей. То же самое можно сказать и о строении сердца. У черепахи формируется неполная перегородка между желудочками, но на более поздних стадиях, чем у цыпленка. Сердце черепахи занимает промежуточное положение между типичным трехкамерным (как у амфибий и ящериц) и четырехкамерным, как у крокодилов и теплокровных.Чтобы подтвердить ведущую роль гена Tbx5в эволюции сердца, были проведены опыты с модифицированными мышами. У этих мышей можно было по желанию экспериментатора отключать ген Tbx5в той или иной части сердечного зачатка. Оказалось, что если выключить ген во всем зачатке желудочков, то зачаток даже не начинает подразделяться на две половинки: из него развивается единый желудочек без всяких следов перегородки. Получаются мышиные зародыши с трехкамерным сердцем! Такие зародыши погибают на 12-й день эмбрионального развития.В другом эксперименте авторам удалось добиться, чтобы ген Tbx5равномерно экспрессировался во всем зачатке желудочков мышиного эмбриона — так же как у лягушки и ящерицы. Это опять-таки привело к развитию мышиных эмбрионов с трехкамерным сердцем.Конечно, было бы еще интереснее сконструировать таких генно-модифицированных ящериц или черепах, у которых Tbx5экспрессировался бы как у мышей и кур, т. е. в левой части желудочка сильно, а в правой — слабо, и посмотреть, не станет ли у них от этого сердце больше похожим на четырехкамерное. Но это пока неосуществимо: генная инженерия рептилий еще не продвинулась так далеко.Ясно, что эволюция для создания теплокровности и всего того, что обеспечивает это преображение (сердце, система кровообращения, покровы, система выделения и т. д.), пользовалась простыми инструментами: чем меньше требуется менять настроек, тем лучше. И если трехкамерное сердце можно превратить в четырехкамерное за один шаг, то нет причин этим не воспользоваться.—————
Назад: Изменения белок-кодирующих генов
Дальше: Дупликация генов

