Книга: Эволюция. Классические идеи в свете новых открытий
Назад: Глава 4 Эволюция на наших глазах
Дальше: Как работает эпистаз
Приспособленность меняется постепенно
Начнем с самого масштабного и самого известного эволюционного эксперимента. В 1988 году группа биологов из Университета штата Мичиган под руководством Ричарда Ленски начала — и продолжает по сей день — уникальный эксперимент на бактериях, который позволил с небывалой доселе детальностью проследить ход эволюции как на уровне генома (накопление мутаций), так и на уровне целого организма (развитие адаптаций).
Эксперимент проводится параллельно с 12 популяциями кишечной палочки Escherichia coli. Эта бактерия — обычный обитатель кишечника теплокровных животных, включая человека. Все 12 подопытных популяций живут в аэробных условиях в жидкой прозрачной среде, где единственной пищей является глюкоза. Именно количество глюкозы является лимитирующим фактором, ограничивающим размножение микробов. С интервалом в сутки из каждой популяции берут небольшую часть (0,1 мл содержимого колбы) и пересаживают в новую колбу с 9,9 мл свежей питательной среды. Там бактерии быстро размножаются, пока не исчерпают запасы глюкозы. Таким образом, в течение каждого суточного цикла начальный период изобилия, когда численность популяции быстро растет, сменяется периодом голода, когда бактерии перестают размножаться и их численность стабилизируется. Численность бактерий оценивают по мутности среды — обычный и весьма надежный метод при наличии соответствующего оборудования.
Периодически часть каждой популяции замораживают при −80 °C (что ничуть не вредит здоровью микробов) и сохраняют для последующего изучения. Это мудро, потому что аналитические методики — в том числе методики секвенирования («прочтения») геномов — сейчас стремительно развиваются и столь же стремительно дешевеют.
Регулярно проводится и оценка приспособленности. Для этого сравнивают скорость размножения подопытных и контрольных бактерий, причем в роли вторых выступают размороженные предки первых — исходный штамм, из которого были взяты родоначальники всех 12 экспериментальных популяций.
Кишечные палочки выгодно отличаются от баобабов тем, что позволяют Ленски и его коллегам ежегодно получать интересные результаты и публиковать статьи в ведущих журналах. Так, в 2009 году они опубликовали в Natureотчет об эволюции одной из 12 популяций ( Barrick et al., 2009). К этому времени длительность эксперимента уже была достаточной для того, чтобы каждая из возможных точечных мутаций (нуклеотидных замен) успела произойти в популяции более одного раза .
Авторы следили за темпом закрепления мутаций и изменением приспособленности. Напомним, что возникновение мутации и ее закрепление — не одно и то же. Далеко не всякая возникшая мутация закрепляется (фиксируется) в популяции. Каждая мутация изначально возникает только у одного микроба. Чтобы мутация зафиксировалась, т. е. достигла 100-процентной частоты, потомки этого микроба должны вытеснить всех остальных микробов в своей колбе. Вредная мутация, скорее всего, будет отсеяна отбором. Полезная мутация под действием отбора может закрепиться, но может и случайно потеряться, пока ее носители еще не успели как следует размножиться. Наконец, нейтральные мутации должны фиксироваться с постоянной скоростью, равной скорости мутирования (обо всем этом мы говорили в главе 1).
За первые 20 тыс. поколений в подопытной популяции зафиксировалось 45 мутаций, в том числе 29 однонуклеотидных замен и 16 иных мутаций (вставок, выпадений, инверсий, встраиваний мобильных элементов). Самое интересное, что скорость накопления мутаций на этом этапе была постоянной. Приспособленность вела себя иначе: сначала она быстро росла, а затем ее рост замедлился (см. рисунок).
Постоянная скорость фиксации, согласно теории, характерна для нейтральных мутаций. Однако все 45 мутаций не могли быть нейтральными. Ясно, что по крайней мере некоторые из них были полезными — об этом свидетельствует рост приспособленности. Полученные результаты трудно увязать и с гипотезой о том, что все 45 мутаций были полезными. Ведь в этом случае обе величины — приспособленность и число накопленных мутаций, — скорее всего, должны были бы меняться сходным образом, т. е. или расти с постоянной скоростью, или параллельно замедляться.
Простейшее объяснение состоит в том, что среди 45 зафиксировавшихся мутаций большинство были нейтральными, а некоторые — полезными. Основная масса полезных мутаций зафиксировалась вскоре после начала эксперимента, т. е. после попадания микробов в новые для них условия, к которым они были плохо приспособлены. Но возможности для «полезного мутирования» быстро исчерпались, и в дальнейшем фиксировались в основном нейтральные мутации.
Есть, однако, четыре аргумента против такого объяснения.
1. В случае преобладания нейтральных мутаций должно быть повышено число синонимичных нуклеотидных замен. Вопреки этим ожиданиям все без исключения зафиксировавшиеся мутации в кодирующих областях генов оказались значимыми (несинонимичными). Резкое преобладание значимых замен над синонимичными — это характернейшая «подпись», оставляемая в геноме положительным отбором.
2. В случае преобладания нейтральных мутаций следует ожидать, что во всех 12 экспериментальных популяциях за 20 тыс. поколений мутации зафиксировались в разныхгенах. Напротив, мутации в одних и тех же генах, закрепившиеся независимо в разных популяциях, будут доводом в пользу того, что мутации фиксировались под действием положительного отбора, а не генетического дрейфа (т. е. мутации были полезными). Чтобы проверить это, авторы отсеквенировали у бактерий поколения № 20 000 из остальных одиннадцати экспериментальных популяций 14 генов, в которых у первой популяции закрепились мутации. Оказалось, что в большинстве случаев в других популяциях эти гены тоже изменились.
3. Если бы большинство мутаций были нейтральными, наблюдалась бы значительная внутрипопуляционная изменчивость по этим локусам (потому что полезные мутации под действием отбора фиксируются быстро, а нейтральные сначала должны долго «случайно блуждать» между нулевой и 100-процентной частотой). Это предположение не подтвердилось.
4. При помощи генной инженерии авторы смогли напрямую определить степень полезности девяти мутаций из рассматриваемых 45. Эти мутации искусственно внедряли в геном предкового штамма. В восьми случаях из девяти приспособленность бактерий резко повысилась . Что касается девятой мутации, то авторы думают, что она тоже полезна, но не сама по себе, а в сочетании с другими мутациями, потому что точно такая же мутация закрепилась у других подопытных популяций.
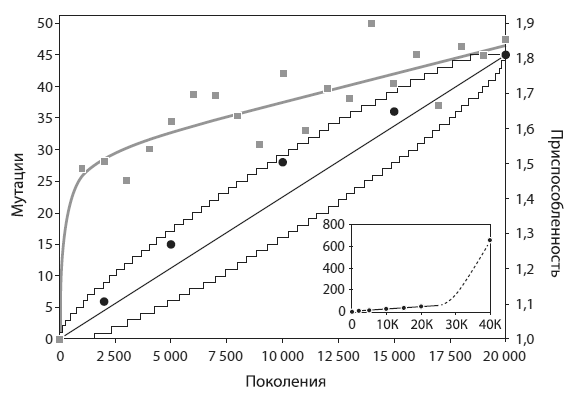 Накопление мутаций(черные линии и кружки) и рост приспособленности(серые линии и квадраты) в экспериментальной популяции. По горизонтальной оси — номер поколения. Видно, что число зафиксировавшихся мутаций росло линейно (тонкие ломаные линии очерчивают 95-процентные доверительные интервалы линейной модели). Приспособленность сначала росла очень быстро, а потом ее рост замедлился. «Прыжки» серых квадратов вверх и вниз относительно серой линии не выходят за пределы статистической погрешности, т. е. не требуют специальных объяснений. На маленьком графике в правом нижнем углу показано резкое ускорение накопления мутаций начиная примерно с поколения № 26 000, когда в популяции зафиксировалась мутация, повышающая темп мутагенеза. ИзBarrick et al., 2009 .
Накопление мутаций(черные линии и кружки) и рост приспособленности(серые линии и квадраты) в экспериментальной популяции. По горизонтальной оси — номер поколения. Видно, что число зафиксировавшихся мутаций росло линейно (тонкие ломаные линии очерчивают 95-процентные доверительные интервалы линейной модели). Приспособленность сначала росла очень быстро, а потом ее рост замедлился. «Прыжки» серых квадратов вверх и вниз относительно серой линии не выходят за пределы статистической погрешности, т. е. не требуют специальных объяснений. На маленьком графике в правом нижнем углу показано резкое ускорение накопления мутаций начиная примерно с поколения № 26 000, когда в популяции зафиксировалась мутация, повышающая темп мутагенеза. ИзBarrick et al., 2009 .
Таким образом, в течение первых 20 тыс. поколений в популяции фиксировались преимущественно полезные мутации, причем их фиксация шла с постоянной скоростью. Замедление роста приспособленности, очевидно, было связано с тем, что средняя степень полезности мутаций постепенно снижалась. Наиболее радикальные адаптивные изменения произошли в течение первых 2 тыс. поколений, а затем происходила более тонкая настройка.
До сих пор речь шла только о первой половине эксперимента. Во второй его половине эволюционная динамика популяции резко изменилась. Дело в том, что после 26 тыс. поколений зафиксировалась мутация в гене mutT. Этот ген кодирует белок, участвующий в репарации (починке) ДНК. В результате частота мутирования выросла примерно в 70 раз (от 1,6×10 −10до 1,1×10 −8на нуклеотид за поколение). Как следствие, более чем на порядок выросла и частота фиксации мутаций. В течение второй половины эксперимента зафиксировалось 609 мутаций — в 13,5 раза больше, чем за первые 20 тыс. поколений.
Аналогичные мутации, увеличившие темп мутагенеза, закрепились и в нескольких других экспериментальных популяциях. Из этого следует, что рост темпов мутагенеза дал бактериям адаптивное преимущество. Это, между прочим, противоречит распространенной идее о том, что в стабильных условиях организмам было бы выгодно снизить темп мутирования до нуля — и этого не происходит только из-за технической невозможности обеспечить абсолютную точность копирования ДНК.
—————МутаторыМутации, повышающие скорость мутирования, закрепляются во многих эволюционных экспериментах на бактериях. Гены (точнее, генетические варианты — аллели), несущие такие мутации, называют аллелями-мутаторами или просто мутаторами. Аллелем-мутатором может стать любой ген, нормальная работа которого важна для точной репликации или репарации, если его слегка «подпортит» случайная мутация.Нужно иметь в виду, что «полезность» мутаторов не прямая, а косвенная. Сама по себе повышенная скорость мутагенеза снижает приспособленность организмов, потому что большинство мутаций, как известно, вредны. Все очень просто: чем интенсивнее мутагенез, тем больше мутаций у потомков и, следовательно, тем ниже их средняяприспособленность. Это и значит, что мутатор не приносит прямой пользы — наоборот, он приносит прямой вред. Косвенная же польза заключается в том, что нет-нет да и возникнет у какого-нибудь носителя аллеля-мутатора редкая, маловероятная полезная мутация. Причем настолько полезная, что ее польза перевесит вред, приносимый мутатором напрямую. Поскольку полезная мутация маловероятна, она скорее возникнет у носителя аллеля-мутатора, чем у организма с низкой скоростью мутирования. В результате адаптивное преимущество получит организм, несущий редкую полезную мутацию, а заодно и аллель-мутатор — в качестве неприятной, но неизбежной «нагрузки». Этот организм и его потомки будут размножаться быстрее других, постепенно вытесняя из популяции конкурентов и распространяя в генофонде обе свои генетические особенности — полезную мутацию и аллель-мутатор. Генетики называют такой способ распространения аллелей hitchhiking— езда автостопом. Имеется в виду, что мутатор «едет автостопом» на полезной мутации, распространяясь в генофонде вопреки тому, что сам по себе он вообще-то вреден.«Автостоп» прекрасно работает в бесполых популяциях, где связку «полезная мутация — аллель-мутатор» невозможно разорвать. Обычно именно с такими бактериями — бесполыми, нарочно лишенными способности к горизонтальному переносу генов — и работают экспериментаторы. В эксперименте Ленски тоже используются бесполые кишечные палочки. Если бы бактерии могли меняться генами, эволюционные судьбы двух аллелей не были бы столь неразрывными. Обязательно появились бы бактерии, несущие полезную мутацию, но не имеющие мутатора. Эти бактерии стали бы вытеснять тех, у кого два аллеля остались «в связке».При действующем горизонтальном переносе генов мутатору куда труднее зафиксироваться. Он может даже быть полностью вытеснен из генофонда — конечно, если не успеет до этого момента сгенерировать у кого-то из своих носителей еще одну полезную мутацию.—————
Мутация, повысившая темп мутагенеза, увеличила вероятность возникновения новых полезных мутаций, когда простые (высоковероятные) пути для этого уже были пройдены. И в этом состояло единственное благо от ускорения мутагенеза. Но при этом в качестве побочного эффекта должно было вырасти число вредных и нейтральных мутаций.
Поэтому следовало ожидать, что теперь большинство фиксирующихсямутаций будут не полезными, а нейтральными. Как мы помним, скорость фиксации нейтральных мутаций в популяции равна скорости мутагенеза. Действительно, в первые 20 тыс. поколений фиксировалось очень мало нейтральных мутаций, а большая часть из 609 «поздних» мутаций оказались нейтральными.
Результаты эксперимента оказались во многом неожиданными. Например, мало кто ожидал, что постоянный темп накопления полезных мутаций может сопровождаться замедляющимся ростом приспособленности или что соотношение темпов фиксации нейтральных и полезных мутаций может так резко меняться. Очевидно, количественные соотношения между разными аспектами эволюционного процесса (нейтральностью и адаптивностью, дрейфом и отбором, темпами изменений на уровне генотипа и фенотипа) могут быть более сложными, неоднозначными и переменчивыми, чем предполагалось.
Назад: Глава 4 Эволюция на наших глазах
Дальше: Как работает эпистаз

