Книга: Эволюция. Классические идеи в свете новых открытий
Назад: Переменчивая среда способствует половому размножению
Дальше: От горизонтального переноса генов к половому размножению
Микробам — горизонтальный перенос, высшим организмам — половое размножение
Рассмотренные примеры говорят о пользе скрещивания и перемешивания генов при половом размножении. Но у бактерий и архей вместо настоящего амфимиксиса работает горизонтальный перенос. Будет ли секс и в этом случае выполнять ту же функцию — быстро приспосабливать организмы к изменившимся условиям? Если это так, то ГПГ у микробов должен быть очень широко распространен и практиковаться не только между популяциями, но главным образом между особями одной популяции. Полезный аллель, который легко позаимствовать, скорее всего, окажется у близкого собрата.
Традиционно преобладала точка зрения, что отбор у бактерий все-таки в основном клональный, действующий на уровне целых геномов. Это значит, что ГПГ не играет существенной роли в повседневных «попытках» прокариот приспособиться к изменчивой среде. Эта гипотеза теперь, при нынешнем развитии биотехнологий, легко проверяема. Такой труд взяли на себя биологи из Массачусетского технологического института: они показали, что традиционная точка зрения неверна или верна только отчасти и что в своей повседневной переменчивой жизни микробы то и дело заимствуют гены у своих ближайших родичей — других микробов той же популяции ( Shapiro et al., 2012).
Биологи работали с двумя популяциями морских планктонных бактерий Vibrio cyclitrophicus. Эти популяции, обозначаемые буквами L и S, находятся в процессе адаптации к разным экологическим нишам: часть бактерий приурочена к крупным (L), а другая — к мелким (S) частицам, отфильтрованным из морской воды. Результаты генетического анализа говорят о том, что разошлись они недавно ( Hunt et al., 2008). По-видимому, популяции L и S приспосабливаются к жизни на разных представителях зоо- или фитопланктона .
Авторы решили выяснить, какой из двух процессов преобладает на начальных этапах экологической дифференциации — отбор отдельных генов с удачными мутациями, которые распространяются за счет ГПГ, или клональный отбор, работающий с целыми геномами.
Различить эти две ситуации можно, сравнив внутри- и межпопуляционный генетический полиморфизм (вариабельность) изучаемых популяций. Если преобладает отбор на уровне генов, две популяции должны четко отличаться друг от друга по небольшому числу генов — тех, от которых зависят экологические свойства популяций. При этом различающиеся участки генома должны иметь пониженный уровень внутрипопуляционного полиморфизма хотя бы в одной из двух популяций. Ведь на каждый такой участок действовал отбор, который поддерживал какой-то один вариант этого участка, вытесняя из генофонда другие его варианты (см. раздел «Следы естественного отбора» в главе 2). Напротив, те участки генома, которые у двух популяций сходны, должны быть более полиморфными в пределах каждой популяции, причем наборы вариантов (аллелей) могут быть одинаковыми в двух популяциях. Ведь отбор, связанный с приспособлением к новой нише, на них не действовал и они могли сохранить исходный полиморфизм, накопленный предками.
Если же в процессе расхождения популяций преобладал клональный отбор, то и уровень полиморфизма, и генетические различия между популяциями должны быть распределены по геному более равномерно.
Авторы отсеквенировали геномы 13 бактерий из популяции L и семи особей из популяции S. Сравнение геномов подтвердило первую версию: в недавней эволюции двух бактериальных популяций явно преобладал отбор на уровне отдельных генов. Следовательно, имел место интенсивный ГПГ между родственными микробами. Вот некоторые факты, на которых основан этот вывод.
Обнаружено 725 нуклеотидных позиций, в которых у микробов L всегда стоит какой-то один нуклеотид, а у микробов S — другой. Эти 725 позиций назвали «экоснипами» (ecoSNPs ), поскольку именно они, скорее всего, отвечают за приспособление к различающимся условиям. Экоснипы не рассеяны по геному хаотически, а сгруппированы в 11 кластеров. Внутри каждого кластера хотя бы у одной из двух популяций наблюдается пониженный уровень полиморфизма, что свидетельствует о недавнем действии положительного отбора.
Все прочие SNP (полиморфные нуклеотидные позиции), в числе 28 744 штук, равномерно рассеяны по геному и варьируют сходным образом и у L-экотипа, и у S-экотипа. Например, в какой-то позиции нуклеотид Ц может стоять у шести, а нуклеотид Г — у семи бактерий L, тогда как в популяции S нуклеотид Ц обнаружен у трех бактерий, а Г — у четырех.
Это соответствует предположению о том, что отбор в сочетании с ГПГ распространял отдельные участки ДНК с удачными мутациями, а не целые геномы. При этом в остальных участках генома сохранялся исходный полиморфизм, имевшийся у предковой популяции.
Другой важный вывод состоит в том, что генетический обмен между популяциями в последнее время происходит редко по сравнению с обменом внутри популяций. В прошлом, когда популяции еще не окончательно разошлись по экологическим нишам, межпопуляционный обмен происходил чаще.
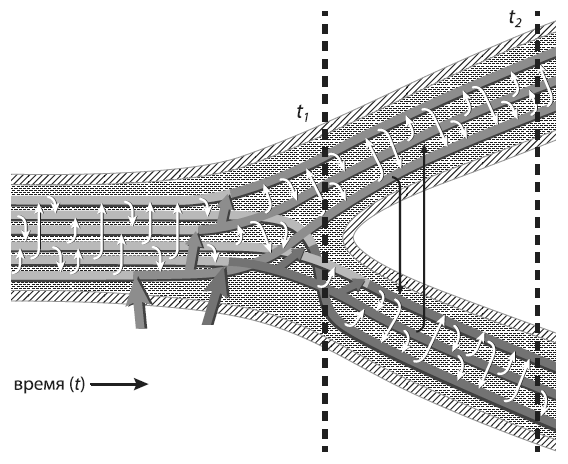 Модель экологической дифференциации двух популяций морской бактерииVibrio cyclitrophicus . Тонкие белые и черные стрелки символизируют внутри- и межпопуляционный ГПГ. Толстые стрелки показывают появление (в результате мутации или ГПГ) адаптивных аллелей, облегчающих жизнь в двух разных местообитаниях. ИзShapiro et al., 2012 .
Модель экологической дифференциации двух популяций морской бактерииVibrio cyclitrophicus . Тонкие белые и черные стрелки символизируют внутри- и межпопуляционный ГПГ. Толстые стрелки показывают появление (в результате мутации или ГПГ) адаптивных аллелей, облегчающих жизнь в двух разных местообитаниях. ИзShapiro et al., 2012 .
Стало быть, пора пересмотреть некоторые устоявшиеся представления. Во-первых, выясняется, что на генетическом уровне эволюционные процессы у прокариот с их ГПГ и эукариот, размножающихся половым путем, различаются не так уж сильно. В обоих случаях отбор идет в основном на уровне отдельных фрагментов ДНК, а не целых геномов.
Во-вторых, стало ясно, что генетический обмен между родственными микробами (представителями одной и той же популяции) идет очень активно, фактически выполняя ту же функцию, что и половой процесс у высших организмов. При этом важнейшую роль играет гомологичная рекомбинация, благодаря которой заимствованные гены не добавляются к уже имеющимся, а заменяют собой свои «старые версии».
В-третьих, высокая частота внутрипопуляционного ГПГ по сравнению с межпопуляционным говорит о том, что у бактерий, приспосабливающихся к разным экологическим нишам, формируется частичная репродуктивная изоляция — совсем как у высших организмов в процессе симпатрического видообразования (см. главу 6). Эта изоляция основана не только на том, что микробы с разными адаптациями живут в разных местах и поэтому редко встречаются, но и на том, что по мере накопления нуклеотидных различий снижается вероятность гомологичной рекомбинации. По-видимому, прокариот все-таки нельзя считать «единым сверхполиморфным видом» (как предлагали некоторые теоретики, исходившие из предположения, что микробы обмениваются генами с кем попало независимо от степени родства). У прокариот, как и у высших организмов, могут существовать частично изолированные популяционные генофонды.
Что же касается случаев ГПГ между неродственными микробами, то они, по-видимому, представляют собой лишь вершину айсберга — легко обнаруживаемый и имеющий важные эволюционные последствия, но все-таки сравнительно редкий побочный эффект способности микробов заимствовать гены у своей ближайшей родни.
Получены и другие данные, указывающие на неожиданно высокие темпы внутривидового ГПГ и гомологичной рекомбинации у бактерий ( Fell et al., 2000; Takuno et al., 2012; Yahara et al., 2012) и архей ( Papke et al., 2004). Конечно, интенсивность генетического обмена у них все равно гораздо ниже той, что характерна для эукариот, практикующих амфимиксис. Главный вывод из этих работ состоит в том, что секс у прокариот играет, по-видимому, ту же роль, что и у эукариот. Он повышает адаптационные возможности организмов путем разрушения связей между полезными и вредными мутациями и позволяет отбору работать с отдельными генами, а не с целыми геномами. Более того, благодаря положительной корреляции между сходством последовательностей ДНК и вероятностью гомологичной рекомбинации (напомним, что это исходный, предковый механизм выбора брачного партнера) ( Majewski et al., 2000; Dubnau, 1999; Thomas, Nielsen, 2005) у микробов формируются хорошо перемешанные видовые генофонды, к которым приложимы классические популяционно-генетические подходы ( Jeltsch, 2003).
Даже родственные микробы, живущие в одних и тех же местообитаниях, могут подразделяться на популяции с полуизолированными генофондами, подобные «биологическим видам» эукариот. Это показано не только для морских бактерий Vibrio, но и для других микробов — например, для совместно встречающихся разновидностей архебактерий Ferroplasma ( Eppley et al., 2007).
Назад: Переменчивая среда способствует половому размножению
Дальше: От горизонтального переноса генов к половому размножению

