Книга: Эволюция. Классические идеи в свете новых открытий
Назад: Вселенная древних белков продолжает расширяться
Дальше: Глава 2 Полезные ошибки
Лабиринт расходящихся тропок
Обычного отрицательного отбора, отсеивающего вредные мутации, явно недостаточно для того, чтобы объяснить медленную эволюцию древних белков. Предположение о том, что каждая аминокислота, стоящая в данной позиции, влияет на приспособленность одинаковым образом независимо от «контекста» (т. е. от того, какие аминокислоты стоят в других позициях в том же белке или в других белках, взаимодействующих с ним), соответствует представлению о легкопроходимомландшафте приспособленности (см. рисунок ниже). Но со времен LUCA в каждом синонимичном сайте (так называют те нуклеотиды в цепи ДНК, изменение которых не влияет на структуру кодируемого белка) произошло уже свыше 100 замен. Синонимичные нуклеотидные замены — это пример движения по ровным горизонтальным поверхностям ландшафта приспособленности. Из этого следует, что в случае хорошей проходимости ландшафта изучаемые белки давным-давно должны были освоить всю потенциально доступную им область в пространстве последовательностей, и тогда мы не наблюдали бы их продолжающегося расхождения.
Резко уменьшить проходимость ландшафта приспособленности — превратить его в лабиринт узких тропок или горных хребтов — может эпистаз. Так называют взаимное влияние разных мутаций (аллелей) на вредность или полезность друг друга. Замена одной аминокислоты может менять эффективность работы белка не сама по себе, а в зависимости от комбинации других аминокислот, занимающих определенные позиции в том же белке. Также эффект мутации может зависеть от других белков, находящихся в непосредственном взаимодействии с белком-мутантом. Иными словами, для определения смысла биологического текста важен контекст. В этом и состоит суть эпистаза. Мы еще не раз встретимся с этим явлением в последующих главах.
Наличие эпистаза предполагает, что многие аминокислотные замены являются допустимыми только в определенном окружении. Если окружение (контекст) неподходящее, то данная замена будет снижать приспособленность, и отбор ее выметет. Это явление в эволюции белков изучалось ранее на конкретных примерах . Если эпистаз широко распространен, белок может добраться до многих потенциально достижимых пунктов в пространстве последовательностей только долгим обходным путем.
Для проверки гипотезы о действенности эпистаза в эволюции белков Поволоцкая и Кондрашов придумали хитроумный тест, основанный на сопоставлении темпов накопления «сближающих» и «разводящих» мутаций с дистанциями между белками. Основная идея состояла в том, что гипотезы о наличии и отсутствии эпистаза дают противоположные предсказания о частоте закрепления сближающих и разводящих мутаций. Если влияние эпистаза невелико, то у двух близких белков темп накопления разводящих мутаций должен быть высоким, а по мере расхождения последовательностей он должен снижаться (если ландшафт приспособленности легкопроходимый, то белки сначала свободно и быстро «блуждают» по ровному плато, соответствующему данной функции, уходя все дальше от исходной точки, но по мере приближения к его краям их расхождение замедляется). Темп накопления сближающих мутаций при этом должен оставаться примерно постоянным. Если же эпистаз оказывает сильное влияние на эволюцию белков, все должно быть наоборот: разводящие мутации должны накапливаться с примерно постоянной скоростью, а темп накопления сближающих мутаций по мере расхождения белков должен снижаться.
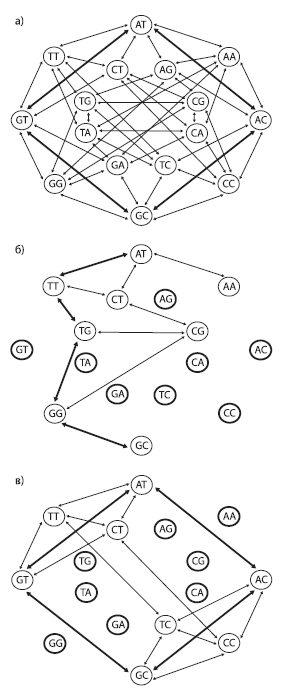 Пространство последовательностей можно изобразить в графа, вершины которого соответствуют разным последовательностям (в данном случае показаны последовательности из двух нуклеотидов), а ребра — единичным эволюционным событиям (нуклеотидным заменам). В первом случае (a) ландшафт приспособленности представляет собой сплошное ровное плато: все 16 последовательностей имеют одинаково высокую приспособленность и все возможные мутации разрешены (не будут отбракованы отбором). Два кратчайших пути, соединяющие последовательности AT и GC, состоят всего из двух мутационных шагов (жирные стрелки). Во втором случае (б) половина последовательностей имеют пониженную приспособленность (отмечены жирными кругами). При этом снижение приспособленности определяется не конкретным нуклеотидом в конкретной позиции, а уникальной комбинацией обоих нуклеотидов (т. е. имеется эпистаз). Это резко снижает проходимость ландшафта: число доступных траекторий уменьшается и кратчайший путь между двумя пунктами (например, от AT к GC) удлиняется. В третьем случае (в) тоже половина последовательностей имеют пониженную приспособленность, но эпистаза нет: нуклеотиды A и G во второй позиции снижают приспособленность независимо от состояния первой позиции. Отсутствие эпистаза способствует тому, что ландшафт остается легкопроходимым, и от AT к GC можно прийти всего за два шага, как и в первом случае. По рисунку изPovolotskaya, Kondrashov, 2010 .
Пространство последовательностей можно изобразить в графа, вершины которого соответствуют разным последовательностям (в данном случае показаны последовательности из двух нуклеотидов), а ребра — единичным эволюционным событиям (нуклеотидным заменам). В первом случае (a) ландшафт приспособленности представляет собой сплошное ровное плато: все 16 последовательностей имеют одинаково высокую приспособленность и все возможные мутации разрешены (не будут отбракованы отбором). Два кратчайших пути, соединяющие последовательности AT и GC, состоят всего из двух мутационных шагов (жирные стрелки). Во втором случае (б) половина последовательностей имеют пониженную приспособленность (отмечены жирными кругами). При этом снижение приспособленности определяется не конкретным нуклеотидом в конкретной позиции, а уникальной комбинацией обоих нуклеотидов (т. е. имеется эпистаз). Это резко снижает проходимость ландшафта: число доступных траекторий уменьшается и кратчайший путь между двумя пунктами (например, от AT к GC) удлиняется. В третьем случае (в) тоже половина последовательностей имеют пониженную приспособленность, но эпистаза нет: нуклеотиды A и G во второй позиции снижают приспособленность независимо от состояния первой позиции. Отсутствие эпистаза способствует тому, что ландшафт остается легкопроходимым, и от AT к GC можно прийти всего за два шага, как и в первом случае. По рисунку изPovolotskaya, Kondrashov, 2010 .
Проведенные расчеты подтвердили гипотезу о сильном влиянии эпистаза на эволюцию белков. Оказалось, что темп накопления разводящих мутаций не зависит от дистанции между белками. В каждый момент времени лишь около 2 % аминокислотных позиций могут быть изменены без снижения приспособленности, хотя в долгосрочной перспективе более 90 % позиций могут измениться — но к этим изменениям нужно идти долгими обходными путями по лабиринту узких «горных хребтов» ландшафта приспособленности. У близких, недавно разошедшихся белков темп накопления сближающих мутаций очень высок, потому что у близких белков аминокислотный «контекст» для каждой позиции является сходным. Поэтому та аминокислота, которая недавно стояла в данной позиции, с большой вероятностью может «вернуться» на свое место, и это не снизит приспособленность. Напротив, у сильно различающихся белков данная позиция уже находится в разных контекстах, и поэтому «возвращение» аминокислоты, которая стояла здесь у далекого предка, скорее всего, снизит приспособленность, и мутация будет отсеяна.
В снижении вероятности сближающих мутаций по мере расхождения белков проявляется правило необратимости эволюции. Чем сильнее разошлись белки, тем меньше у них шансов снова стать похожими (мы еще вернемся к этому правилу в главе 5).
Таким образом, ландшафт приспособленности древних белков отличается повышенной «складчатостью». Он похож на сложный лабиринт узких горных хребтов, передвигаться по которому эволюционирующие белки могут лишь очень медленно. Мало того, некоторые узкие мостики норовят обрушиться в бездну после того, как по ним «прополз» эволюционирующий белок, — совсем как в голливудских боевиках, — что придает эволюции элемент необратимости.
Между прочим, полученные результаты — еще один аргумент в пользу единства происхождения всего живого. Подобно тому как разбегающиеся галактики указывают на существовавший в прошлом единый центр, из которого все объекты во Вселенной начали свое движение, так и продолжающееся «разбегание» древних белков указывает на их происхождение от единого предка.

