Книга: Самое грандиозное шоу на Земле: доказательства эволюции
Назад: ГЛАВА 7 ОТСУТСТВУЮЩИЕ ОСОБИ? БОЛЬШЕ НЕ ОТСУТСТВУЮЩИЕ
Дальше: ГЛАВА 9 КОВЧЕГ КОНТИНЕНТОВ
ГЛАВА 8 ВЫ СДЕЛАЛИ ЭТО ЗА ДЕВЯТЬ МЕСЯЦЕВ САМИ
ЭТОТ вспыльчив гений Дж. Б. С. Холдейн, который сделал так много помимо того, что был одним из трех ведущих архитекторов неодарвинизма, однажды получил вызов от леди после общественной лекции. Есть устная история, и Джон Мейнард Смит, к сожалению, не может подтвердить точность слов, но разговор был приблизительно таким:
Скептик эволюции: профессор Холдейн, даже с учетом миллиардов лет, которые, как вы говорите, были доступны для эволюции, я просто не могу поверить, что можно перейти от одной клетки к сложным человеческим телам, с их триллионами клеток, организованных в кости и мускулы, и нервы, сердце, которое качает, не прекращая, в течение многих десятилетий, мили и мили кровеносных сосудов и почечных трубочек, и мозг, способный к размышлению, и разговору, и чувству.
Дж. Б. С. Холдейн: Но мадам, вы сделали это сами. И вам потребовались только девять месяцев.
Спросившая была, возможно, на мгновение ошарашена неожиданностью ответа Холдена. Выбить почву из-под ног -показалось бы преуменьшением. Но , возможно, возражение Холдейна оставило ее без удовлетворения. Я не знаю, спросила ли она дополнительно, но, если это так, это могло бы идти в таком направлении:
"Скептик эволюции: Ах да, но развивающийся эмбрион следует генетическим инструкциям. Эта инструкция по созданию сложного тела, как вы, профессор Холдейн, утверждаете, эволюционировала путем естественного отбора. И мне до сих пор трудно поверить, даже с учетом миллиардов лет, предоставленных для эволюции.
Возможно, она была права. И даже если божественный разум действительно, в конечном счете, окажется ответствен за проектирование сложности живого, определенно не верно, что он вылепливает живые организмы в форме глиняных или подобных моделей, в которых плотники, гончары, портные или производители автомобилей решают свои задачи. Мы можем быть "чудесно развитыми", но не "чудесно сделанными". Когда дети поют, "Он сделал их яркие краски / Он сделал их крошечные крылья", они произносят по-детски очевидную неправду. Чем бы ни занимался Бог еще, он, конечно, не делает ярких красок и крошечные крылья. Если бы он вообще делал что-нибудь, это должно было бы контролировать эмбриональное развитие существ, например, сшивая вместе последовательность генов, которые направляют процесс автоматизированного развития. Крылья не сделаны, они растут -постепенно - из почек конечностей в яйце.
Бог, повторю этот важный момент, который должен был бы быть очевидным, но не очевиден, никогда не делал крошечное крыло за всю свою вечную жизнь. Если он и сделал что-нибудь (по моему мнению, он не делал, но давайте это пропустим; это не то, о чем я веду речь здесь), то, что он сделал, был эмбриологический рецепт или что-то вроде компьютерной программы для управления эмбриональным развитием крошечного крыла (плюс также многих других вещей). Конечно, Бог мог бы претендовать на то, что разработать рецепт или программу для крыла - это столь же умно, точно такое же захватывающее дух достижение мастерства, как и сделать крыло. Но в данный момент я лишь хочу рассмотреть различие между созданием чего-то подобного крылу, и тем, что действительно происходит в эмбриологии.
БЕЗ ХОРЕОГРАФА
Ранняя история эмбриологии была расколота на две противостоящие доктрины, названные преформизм и эпигенез. Различие между ними не всегда четко понимают, поэтому я должен потратить немного времени, объясняя эти два термина. Преформисты полагали, что яйцеклетка (или сперматозоид, поскольку преформисты подразделялись на "овистов" и "спермистов") содержит крошечного миниатюрного младенца или "гомункулуса". Все части младенца точно размещались на своих местах, правильно расположенные друг относительно друга, ожидая только раздувания, подобно поделенному на секции воздушному шару. Это вызывает очевидные проблемы. Во-первых, по крайней мере в своей ранней наивной форме, это подразумевает то, что как всем известно, является неправдой: что мы наследуем только от одного родителя - от матери у овистов, от отца у спермистов. Во-вторых, преформисты такого рода должны были столкнуться с проблемой бесконечного регресса в стиле матрешки из гомункулов внутри гомункулов - или, если не бесконечно, то, по крайней мере, достаточно долго, чтобы довести нас назад к Еве (Адаму у спермистов). Единственным спасением от регресса будет построение гомункула заново в каждом поколении через искусное сканирование взрослого тела предыдущего поколения. Этого "наследования приобретенных признаков" не происходит - иначе еврейские мальчики рождались бы без крайней плоти, а часто посещающие спортзал культуристы (но не их близнецы-диванные овощи) зачинали бы младенцев с кубиками пресса, плеч и ягодиц.
Чтобы быть справедливым к преформистам, они действительно встречали, честно и прямо, эту логическую необходимость регресса, как бы абсурдно это казалось. По крайней мере некоторые из них действительно полагали, что первая женщина (или мужчина) содержала миниатюрных эмбрионов всех ее потомков, вложенных друг в друга, как матрешки. И в некотором смысле они должны были верить в это: смысл, который стоит отметить, поскольку он предвосхищает суть этой главы. Если Вы уверены, что Адам был "сделан", а не родился, вы подразумеваете, что Адам не имел генов -или по крайней мере не нуждался в них, чтобы развиться. Адам не имел никакой эмбриологии, но просто возник. Смежный логический вывод привел Викторианского автора Филипа Госса (отца в "Отец и сын" Эдмунда Госса) к написанию книги под названием "Омфал" (от греческого "пуп") утверждавшей, что у Адама, должен был быть пуп, даже при том, что он никогда не был рожден. Более сложным следствием "омфалогических" рассуждений было бы то, что звезды, расстояние до которых от нас - больше чем несколько тысяч световых лет, должно быть, были созданы с готовыми лучами света, простирающимися почти всю дорогу к нам - иначе мы не смогли бы увидеть их до момента в далеком будущем! Высмеивание омфалогии кажется легкомысленным, существует серьезный момент в эмбриологии, который и является предметом этой главы. Это довольно сложный для понимания момент - на самом деле, я сам в процессе постижения - и я приближаюсь к нему с различных сторон.
По изложенным причинам, преформизм, по крайней мере в его оригинальной версии "матрешки", всегда был обречен на неудачу. Существует ли вариант преформизма, в котором он смог бы возродиться сегодня, в эпоху ДНК? Возможно, но я сомневаюсь, что сможет. Учебники биологии повторяют снова и снова, что ДНК -"чертёж" для строительства тела. Это не так. Настоящий чертёж, скажем, автомобиля или дома воплощает соответствие один-к-одному бумаги и готового изделия. Из этого следует, что чертёж является обратимым. Столь же легко перейти от дома к чертежу, как и наоборот, именно потому, что это соответствие один-к-одному. На самом деле это еще проще, поскольку дом надо строить, но вам надо сделать только несколько замеров, чтобы затем нарисовать чертёж. Если вы возьмете тело животного, не важно сколько подробных замеров вы произведете, вы не сможете восстановить его ДНК. Это то, что делает ложным высказывание, что ДНК представляет собой чертёж.
Теоретически можно вообразить - возможно, это происходит на некоторой чужой планете - что ДНК может быть закодированным описанием тела: своего рода трехмерной картой, представленной в линейном коде "букв" ДНК. Так оно действительно было бы обратимо. Сканирование тела, чтобы сделать генетический чертёж, не является совершенно нелепой идеей. Если бы ДНК работала так, мы могли бы представить это как своего рода нео-преформизм. И это не вызывало бы призрак матрешек. Мне не ясно, вызывало ли бы это призрак наследования только от одного родителя. У ДНК есть потрясающе точный способ объединять половину отцовской информации с ровно половиной материнской информации, но как бы это происходило с соединением половины результатов сканирования тела матери и половины результатов сканирования тела отца? Не обращайте внимания: все это очень далеко от реальности.
Итак, ДНК решительно не является чертежом. В отличие от Адама, который был вылеплен непосредственно в его взрослой форме, все реальные тела развиваются и растут из единственной клетки через промежуточные стадии эмбриона, плода, младенца, ребенка и подростка. Возможно, в некотором ином мире живущие там существа собирают себя от макушек до пальцев ног, как упорядоченное множество трехмерных био-пикселей, вычитанных из закодированных сканирующих строк. Но это не тот способ, который работает на нашей планете, и на самом деле я думаю, что есть причины, которые я излагал в другом месте (и поэтому здесь не буду вдаваться в подробности), почему это невозможно ни на какой планете.
Исторической альтернативой преформизму является эпигенез. Если преформизм полностью связан с чертежами, то суть эпигенеза заключается скорее в чем-то похожем на рецепт или компьютерную программу. Определение Малого оксфордского английского словаря довольно современно, и я не уверен, что Аристотель, который выдумал слово, узнал бы его: эпигенез: теория развития организма через прогрессивное дифференцирование первоначально недифференцированного целого.
"Принципы развития" Льюиса Вольперта с коллегами описывает эпигенез как идею, что новые структуры возникают последовательно. Есть смысл, при котором эпигенез самоочевидно верен, но детали важны, и в этом клише кроется дьявол. Как организм может развиваться последовательно? Откуда первоначально недифференцированное целое "знает", как дифференцироваться постепенно, если не следуя чертежу? Различие, которое я хочу описать в этой главе, в значительной степени соответствует различию между преформизмом и эпигенезом, это различие между плановой архитектурой и самосборкой. Смысл плановой архитектуры для нас ясен, потому что мы видим ее вокруг себя в наших зданиях и других артефактах. Самосборка менее знакома и будет нуждаться в некотором пояснении. В области биологии развития самосборка занимает позицию, аналогичную естественному отбору в эволюции, хотя она и не является тем же процессом. Обе автоматическими, непреднамеренными, не спланированными средствами достигают результатов, выглядящих, при поверхностном взгляде, как-будто они были детально спланированы.
Дж. Холдейн высказал леди, задавшей вопрос, простую истину, но он не стал бы отрицать, что существует загадка, граничащая с чудом (но никогда вполне не становящаяся им) в том факте, что единственная клетка дает начало человеческому телу во всей его сложности. И загадка только немного проясняется тем, что это достигается с помощью инструкций ДНК. Причина, по которой сохраняется загадка, состоит в том, что нам трудно вообразить даже в принципе, как мы могли бы приступить к написанию инструкции для строительства тела тем способом, каким оно фактически построено, а именно к тому, что я только что назвал "самосборкой", связанной с тем, что программисты иногда называют "восходящей" процедурой, в отличие от "нисходящей".
Архитектор проектирует великий храм. Затем, через иерархическую цепь команд, строительная операция разбивается на отдельные участки, которые разбивают ее дальше на под-участки, и так далее, пока, наконец, инструкции не розданы отдельным каменщикам, плотникам и стекольщикам, которые идут работать, пока не построен храм, выглядящий в значительной мере похожим на исходный чертеж архитектора. Это - нисходящее проектирование.
Восходящее проектирование работает совершенно иначе. Я никогда не верил этому, но существовал миф, что у некоторых из самых прекрасных средневековых соборов в Европе не было архитектора. Никто не проектировал собор. Каждый каменщик и плотник занимался, в соответствии со своей квалификацией, своим собственным небольшим уголком здания, обращая мало внимания на то, что делают другие, и не обращая внимания на какой-либо общий план. Так или иначе, из такой анархии, появлялся собор. Если так действительно происходило, это была восходящая архитектура. Если не брать в расчет миф, то, конечно, это не то, что происходило в случаях с соборами. Но это в значительной степени то, что происходит при строительстве термитника или муравейника - и при развитии эмбриона. Это - то, что делает эмбриологию настолько удивительно отличной от чего-либо, с чем мы, люди, знакомы в отношении строительства или производства.
Тот же принцип работает с определенными типами компьютерных программ, определенными типами поведения животных, и - сводя их воедино - с компьютерными программами, разработанными для моделирования поведения животных Предположим, мы хотим понять стайное поведение скворцов. Существует несколько потрясающих фильмов доступных на YouTube, кадры из которых представлены на цветной странице 16. Эти балетные маневры были сфотографированы возле Отмура, поблизости Оксфорда, Диланом Винтером. Что поразительно в поведении скворцов, это то, что, несмотря на всю видимость, у них нет ни хореографа, ни, насколько нам известно, лидера. Каждая отдельная птица только следует локальным правилам.
Количество отдельных птиц в этих скоплениях может достигать тысяч, и все же они практически никогда не сталкиваются. Это только к лучшему, так как, учитывая скорость, с которой они летают, любое такое столкновение серьезно бы их ранило. Часто целое скопление, кажется, ведет себя как единый индивид, сворачивающий и описывающий виражи. Может выглядеть, как будто отдельные стаи движутся друг сквозь друга в противоположных направлениях, сохраняя нетронутым свое единство как отдельных стай. Это выглядит почти как чудо, но на самом деле стаи находятся на различных расстояниях от камеры и не буквально пролетают друг сквозь друга. Добавляет эстетическое удовольствие то, что края скоплений так резко очерчены. Они не плавно сходят на нет, а имеют резкую границу. Плотность птиц на границе не меньше, чем в середине скопления, в то время как она равна нулю за пределами границы. Как только вы подумаете об этом таким образом, разве это не крайне удивительно?
Все это могло бы послужить более чем элегантной экранной заставкой на компьютере. Вам бы не захотелось реального фильма скворцов, поскольку Ваш скринсейвер повторял бы одни и те же идентичные балетные движения снова и снова, и поэтому не нагружал бы все пиксели одинаково. То, чего вы хотели бы, это компьютерное моделирование стай скворцов; и, любой программист скажет Вам, есть правильный и неправильный способ сделать это. Не пытайтесь поставить весь балет, это было бы ужасно плохим стилем программирования для такого рода задач. Следует поговорить о лучшем способе сделать это, потому что подобным образом, почти наверняка, запрограммированы сами птицы, в их мозгах. Главное -это большая аналогия с тем, как работает эмбриология.
Вот как следует программировать стайное поведение у скворцов. Посвятите почти все ваши усилия программированию поведения одной отдельной птицы. Встройте в вашего робо-скворца детализированные правила того, как лететь и как реагировать на присутствие соседних скворцов, в зависимости от их расстояния и относительного положения. Заложите в правилах, какую важность придавать поведению соседей и какую важность уделить индивидуальной инициативе в изменении направления. Этим типовым правилам помогли бы тщательные наблюдения реальных птиц в действии. Наделите свою киберптицу определенной тенденцией чередовать свои правила в случайном порядке. Теперь, когда вы написали сложную программу, чтобы задать правила поведения одного скворца, наступает определяющий момент, который я подчеркиваю в этой главе. Не пытайтесь программировать поведение целой стаи, как, возможно, делало предыдущее поколение программистов. Вместо этого клонируйте того одного компьютерного скворца, которого вы запрограммировали. Сделайте тысячу копий своей робо-птицы, возможно, одинаковых, или, возможно, с некоторыми небольшими случайными различиями в правилах поведения. И теперь "выпустите" тысячи моделей скворцов в вашем компьютере, так чтобы они могли свободно взаимодействовать друг с другом, подчиняясь все тем же правилам.
Если у вас будут поведенческие правила для одного скворца, то тысяча компьютерных скворцов, каждый из которых - точка на экране, будут вести себя как реальные скворцы, собирающиеся в стаи зимой. Если стайное поведение не вполне правильно, Вы можете вернуться и поправить поведение отдельного скворца, возможно, в свете дальнейших исследований поведения реальных скворцов. Теперь клонируйте новый вариант тысячу раз вместо той тысячи, которая работала не безупречно. Продолжайте переделывать Вашу программу одного клонированного скворца, пока стайное поведение тысяч их на экране не будет убедительной, реалистичной заставкой. Крейг Рейнольдс написал программу по типу этой (не конкретно для скворцов) в 1986 году, назвав ее "Boids".
Ключевым моментом является то, что нет никакого хореографа и никакого лидера. Порядок, организация, структура - все это появляется как побочный продукт правил, соблюдаемых локально и много раз, не глобально. И именно так работает эмбриология. Все делается по локальным правилам на различных уровнях, но особенно на уровне отдельной клетки. Нет хореографа. Нет дирижера у этого оркестра. Нет централизованного планирования. Нет архитектора. В области биологии развития или в производстве эквивалентом этого вида программирования является самосборка.
Тело человека, орла, крота, дельфина, гепарда, лягушки леопарда, ласточки: все они так красиво собраны, кажется, невозможно поверить, что гены, программирующие их развитие, функционируют не как чертеж, проект, генеральный план. Но нет: как и с компьютерными скворцами, все это делается отдельными клетками, подчиняющимися локальным правилам. Красиво "спроектированное" тело появляется как следствие правил, локально выполняемых отдельными клетками, без обращения к чему-либо, что можно было бы назвать всеобщим глобальным планом. Клетки развивающегося эмбриона поворачиваются и танцуют друг вокруг друга, как скворцы в гигантских стаях. Есть отличия, и они важны. В отличие от скворцов, клетки физически соединены друг с другом в пласты и блоки: их "стаи" называют "тканями". Когда они вертятся и танцуют как миниатюрные скворцы, следствием является то, что формируются трехмерные формы, поскольку ткани впячиваются в ответ на движения клеток; или раздуваются или сморщиваются из-за локальных особенностей роста и смерти клеток. Мне нравится аналогия этого в виде искусства оригами, предложенная выдающимся эмбриологом Льюисом Уолпертом в его книге "Триумф эмбриона"; но прежде чем перейти к ней, я должен убрать с пути некоторые альтернативные аналогии, которые могли бы прийти на ум - аналогии из числа человеческих ремесел и производственных процессов.
АНАЛОГИИ ДЛЯ РАЗВИТИЯ
Удивительно трудно найти хорошую аналогию для развития живой ткани, но Вы можете найти частичные сходства с определенными аспектами процесса. Рецепт отражает часть истины, и это - аналогия, которую я иногда использую, чтобы объяснить, почему "чертёж" неуместен. В отличие от чертежа, рецепт необратим. Если Вы будете следовать рецепту пирога шаг за шагом, то Вы получите пирог. Но Вы не можете взять пирог и восстановить рецепт - определенно, не точные слова рецепта - хотя, как мы видели, Вы можете взять дом и восстановить что-то близкое к оригинальному чертежу. Причина в соответствии один-к-одному между частями дома и частями чертежа. С очевидными исключениями, такими как вишенка сверху, нет никакого соответствия один-к-одному между частями пирога и словами, скажем, или предложениями из его рецепта.
Какие могут быть другие аналогии с человеческим производством? Скульптура - в основном полностью мимо цели. Скульптор начинает с куска камня или дерева и придает ему форму удалением, откалывая до тех пор, пока все, что осталось, не будет требуемой формой. Существует, признаю, довольно точное подобие одному особому процессу в эмбриологии, называемому апоптоз. Апоптоз -запрограммированная клеточная смерть, и он задействован, например, в развитии пальцев рук и ног. В человеческом эмбрионе все пальцы рук и ног объединены. В матке Вы и я имели перепончатые ноги и руки. Перепонки исчезли (у большинства людей: есть редкие исключения) из-за запрограммированной смерти клеток. Это немного напоминает способ, которым скульптор вырезает форму, но он не столь распространен или достаточно важен, чтобы отражать то, как обычно работает эмбриология. Эмбриологи могут ненадолго вспомнить о "ваянии скульптора", но они не позволят этой мысли надолго задерживаться.
Некоторые скульпторы работают, не удаляя высеканием, а взяв ком глины или мягкий воск, и вылепливая в форму (которая может впоследствии быть отлита, в бронзе, например). Это опять-таки не удачная аналогия для эмбриологии. Также неудачной является искусство покроя или пошива одежды. Уже существующие ткани разрезаются, формируя расположение по заранее спланированному шаблону, затем сшиваются с другими вырезанными формами. Они часто потом выворачиваются наизнанку, чтобы скрыть швы - и это неточная но, по крайней мере, хорошая аналогия с определенными частями эмбриологии. Но в целом, эмбриология не более похожа на пошив, чем на ваяние скульптуры. Вязание может быть лучше в том смысле, что все формы свитеров, скажем, строятся из многочисленных отдельных стежков, как отдельных клеток. Но есть лучшие аналогии, как мы увидим.
Как насчет сборки автомобиля, или другой сложной машины на заводском конвейере: это - хорошая аналогия? Как ваяние и пошив, монтаж готовых деталей - эффективный способ сделать что-нибудь. На автомобильном заводе части имеются уже изготовленными предварительно, часто путем отливки в формы в литейном цехе (и есть, я думаю, нечто отдаленно напоминающее литье в эмбриологии). Затем готовые части соединяются на конвейере и привинчиваются, приковываются, свариваются или склеиваются, шаг за шагом, согласно точно начерченному чертежу. Еще раз, у эмбриологии нет ничего напоминающего ранее начерченный чертёж. Но есть сходство с упорядоченным склеиванием предварительно собранных частей, как в сборочном цехе автомобильного завода ранее произведенные карбюраторы, и распределительные головки, и ремни вентилятора, и головки цилиндра сводятся воедино и соединяются в правильном положении.
Ниже приведены три вида вируса. Слева - вирус табачной мозаики (ВТМ), которая паразитирует на растениях табака и других представителях семейства пасленовых, таких, как помидоры. В середине аденовирус, который поражает дыхательную систему многих животных, включая нас. Справа бактериофаг T4, который паразитирует на бактерии. Он похож на лунный посадочный модуль, и он ведет себя довольно схоже, "приземляясь" на поверхность бактерии (которая намного больше), потом становясь на свои "паучьи ноги" , затем проталкивая зонд в середине сквозь клеточную стенку бактерии и впрыскивая внутрь свою ДНК. Вирусная ДНК затем захватывает оборудование бактерии, производящее белки, которое перенастраивается для создания новых вирусов. Другие два вируса на картинке делают что-то подобное, хотя они не выглядят или не ведут себя как лунные аппараты. Во всех случаях их генетический материал захватывает производящий белки аппарат клетки-хозяина и переключает ее молекулярную производственную линию для изготовления в большом количестве вирусов вместо ее нормальных продуктов.
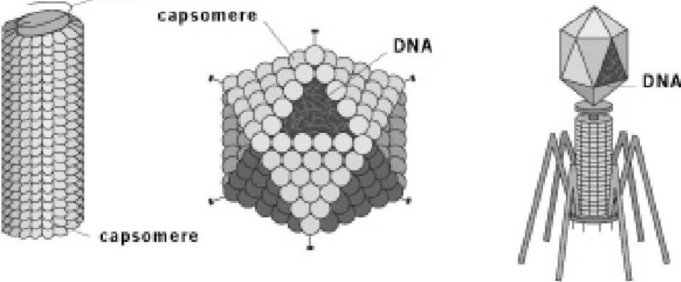
Три вида вируса
Большая часть того, что вы видите на картинках, является белковым контейнером для генетического материала, а в случае Т4 ("лунного посадочного модуля") - оборудованием для заражения хозяина. Интересен способ, которым этот белковый аппарат собирается. Он действительно самосборный. Каждый вирус собирается из нескольких ранее произведенных молекул белка. Каждая молекула белка, способом, который мы увидим, ранее самособралась в характерную "третичную структуру" в соответствии с законами химии, с учетом его конкретной последовательности аминокислот. И затем в вирусе молекулы белка соединяются друг с другом, чтобы сформировать так называемую "четвертичную структуру", снова следуя локальным правилам. Не существует никакого глобального плана, никакого чертежа.
Субблоки белка, которые соединяются как кирпичи Lego, в форме четвертичной структуры, называют капсомерами. Обратите внимание, как геометрически совершенны эти маленькие конструкции. Аденовирус в середине имеет ровно 252 капсомера, нарисованные здесь как маленькие шарики, расположенные в виде икосаэдра. Икосаэдр является платоновской совершенной объемной фигурой, которая имеет 20 треугольных граней. Капсомеры расположены икосаэдром не по какому-либо генеральному плану или чертежу, а просто каждый из них локально повинуется законам химического притяжения, когда наталкивается на других подобных ему. Это тоже, что и формирование кристаллов, и, по сути, аденовирус может быть описан как очень маленький полый кристалл. "Кристаллизация" вирусов - особенно красивый пример "самосборки", который я рекламирую как главный принцип, посредством чего собираются живые существа. "Лунный посадочный модуль", фаг Т4 также имеет икосаэдр в своем основном хранилище ДНК, но его самоорганизующаяся четвертичная структура является более сложной, включающей дополнительные единицы белка, собранные в соответствии с другими локальными правилами в аппарате инъекции и "ногах", прикрепленных к икосаэдру.
Возвращаясь от вирусов к эмбриологии более крупных существ, я прихожу к моей любимой аналогии с человеческой техникой конструирования: оригами. Оригами - это искусство конструктивного сворачивания бумаги, развитого на наиболее продвинутом уровне в Японии. Единственным оригами, которое я знаю как собрать, является "Китайская джонка". Я выучился этому от моего отца, изучавшему его при повальном увлечении, которое прокатилось по его школе в 1920-ых. Одна биологически реалистичная особенность состоит в том, что " эмбриология" китайской джонки проходит через несколько промежуточных "личиночных" стадий, которые сами по себе являются привлекательными творениями, так же как гусеница -красивое, работоспособное промежуточное звено на пути к бабочке, которую она едва напоминает вообще. Начав с простого квадратного листка бумаги, и просто сворачивая его - никогда не разрезая, никогда не склеивая и никогда не внося какие-либо другие части, процедура проходит у нас через три узнаваемых "личиночных стадии": "катамаран", "ящик с двумя крышками "и "картину в раме" до достижения кульминации во "взрослой" китайской джонке. В пользу аналогии с оригами, когда вас впервые учат, как сделать китайскую джонку, не только сама джонка, но и каждая из трех "личиночных" стадий - катамаран, ящик, рамка картины - возникает неожиданно. Ваши руки могут это складывать, но Вы решительно не следуете чертежу китайской джонки или любой из личиночных стадий. Вы следуете ряду правил складывания, у которых, кажется, нет никакой связи с конечным продуктом, пока он наконец не появляется как бабочка из своей куколки. Таким образом, аналогия оригами отражает нечто вроде важности "локальных правил", в отличие от глобального плана.
Также в пользу аналогии оригами, складывание, сворачивание и выворачивание наизнанку являются частью излюбленных уловок, используемых эмбриональными тканями при создании тела. Аналогия особенно хорошо работает на ранней эмбриональной стадии. Но она имеет свои недостатки, и вот два очевидных. Во-первых, для складывания необходимы человеческие руки. Во-вторых, развивающийся бумажный "эмбрион" не становится больше. Он заканчивает, веся ровно столько, как и вначале. Чтобы удостоверить различие, я буду иногда упоминать биологическую эмбриологию как "раздувающееся оригами", а не просто "оригами".
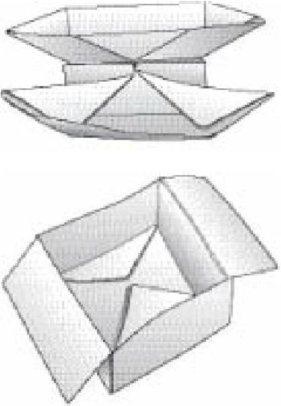
Китайское оригами с тремя "личиночными стадиями»: "катамаран", "коробка с двумя крышками" и "картинка в рамке"
Фактически, эти два недостатка отчасти уравновешивают друг друга. Листки тканей, сгибающиеся, сворачивающиеся и выворачивающиеся наизнанку в развивающемся эмбрионе, действительно растут, и это тот самый рост, обеспечивающий часть движущей силы, которая в оригами вводится человеческой рукой. Если вы хотите сделать модель оригами из листа живой ткани вместо мертвой бумаги, существует, по крайней мере, существенный шанс, что, если лист будет расти только правильным способом, не однородно, а быстрее в некоторых частях листа, чем в других, то это может автоматически заставить лист принимать определенную форму - и даже складываться, сворачиваться или выворачиваться наизнанку определенным способом - не требуя рук для деформации и складывания, и не требуя какого-либо глобального плана, а лишь локальных правил. И на самом деле это больше, чем просто существенный шанс, поскольку это действительно происходит. Давайте назовем это "автооригами". Как практически автооригами работает в эмбриологии? Оно работает, потому что то, что происходит в реальном эмбрионе, когда растет слой ткани - это деление клеток. И неодинаковый рост различных частей слоя ткани в каждой части слоя достигается клетками, делящимися со скоростью, определяемой локальными правилами. Так, окольными путями, мы возвращаемся к фундаментальной важности превосходства восходящих снизу-вверх локальных правил, в противоположность нисходящим сверху-вниз глобальным правилам. Существует целый ряд (гораздо более сложных) версий этого простого принципа, на самом деле происходящих на ранних стадиях эмбрионального развития.
Вот как работает оригами на ранних стадиях развития позвоночных. Единственная оплодотворённая яйцеклетка делится, создавая две клетки. Затем эти две делятся, создавая четыре. И так далее, с быстро удваивающимся и удваивающимся числом клеток. На данном этапе нет никакого роста, никакого раздувания. Первоначальный объем оплодотворённой яйцеклетки буквально разделен на части, как при разрезании пирога, и мы заканчиваем сферическим шаром клеток, имеющим те же размеры, что и первоначальное яйцо. Это не сплошной шар, а полый, и его называют бластулой. Следующая стадия, гаструляция, является предметом известного остроумного выражения Льюиса Уолперта: "Не рождение, свадьба, или смерть, а гаструляция является действительно самым важным временем в Вашей жизни". Гаструляция - своего рода микрокосмическое землетрясение, которое несется по поверхности бластулы и полностью изменяет всю ее форму. Ткани эмбриона сильно перестраиваются.
Гаструляция обычно подразумевает вдавливание полого шара, являющегося бластулой, так, что он становится двухслойным с отверстием во внешний мир (см. стр. с компьютерной симуляцией). Внешний слой этой "гаструлы" называют эктодермой, внутренний слой - эндодермой, и есть также некоторые клетки, попадающие в пространстве между эктодермой и эндодермой, которые называют мезодермой. Каждый из этих трех первичных слоев предназначен для создания крупнейших частей тела. Например, внешняя кожа и нервная система образуются из эктодермы; кишечник и другие внутренние органы образуются из эндодермы; а мезодерма обеспечивает мышцы и кости.
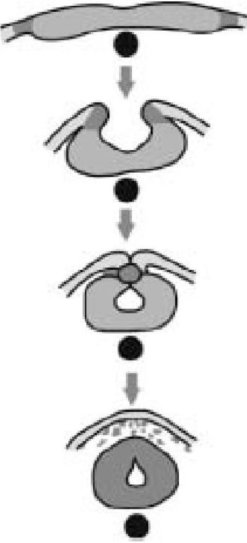
Neurulation
Следующую стадию в оригами эмбриона называют нейруляцией. На схеме справа показано поперечное сечение середины спины нейрулирующего эмбриона земноводного (это могла быть или лягушка, или саламандра). Черный круг - "нотохорда", придающий жесткость стержень, который действует как предшественник позвоночника. Нотохорда является отличительной особенностью типа хордовых, к которому принадлежим мы и все позвоночные животные (хотя у нас, как и у большинства современных позвоночных животных, она есть только у эмбрионов). В нейруляции, как и в гаструляции, инвагинация [впячивание] очень показательна. Вы помните, я говорил, что нервная система происходит из эктодермы. Ну так вот как это происходит. Участок эктодермы впячивается (все дальше назад вдоль тела как застежка-молния), свертывается в трубку, и стягивается там, где стороны трубки "застегиваются на молнию", так что оказывается проходящим вдоль всего тела между внешним слоем и нотохордой. Этой трубке предстоит стать спинным мозгом, главным нервным стволом тела. Передний конец ее набухает и становится мозгом. А все остальные нервы получаются из этой первичной трубки благодаря последующему делению клеток.
Я не хочу вдаваться в детали гаструляции или нейруляции, только скажу, что они замечательны, и что метафора оригами вполне прилично сохраняется в обеих. Я рассмотрел общие принципы, благодаря которым эмбрионы становятся более сложными с помощью раздувающегося оригами. Ниже приведена одна из тех вещей, которую слои клеток, как было подмечено, делают в течение эмбрионального развития, например во время гаструляции. Вы можете легко увидеть, как такая инвагинация могла быть полезным действием для раздувающегося оригами, и она действительно играет крупнейшую роль и в гаструляции, и в нейруляции.

Внедрение в листе клеток
Гаструляция и нейруляция завершаются в раннем развитии, и они затрагивают всю форму эмбриона. Инвагинация и другие маневры "раздувающегося оригами" обеспечивают эти стадии ранней эмбриологии, и они, и подобные уловки также участвуют позднее в развитии, когда создаются специализированные органы, такие как глаза и сердце. Но, если нет рук для сворачивания, какой механический процесс обеспечивает эти динамичные перемещения? Отчасти, как я уже сказал, простое расширение. Клетки размножаются по всему слою ткани. Его площадь, следовательно, увеличивается и, за неимением пространства, у него мало выбора, кроме как скручиваться или впячиваться. Но этот процесс более управляем, и он был расшифрован группой ученых, объединенных блестящим математическим биологом Джорджем Остером из Калифорнийского университета в Беркли.
МОДЕЛИРОВАНИЕ КЛЕТОК ПОДОБНО СКВОРЦАМ
Остер и его коллеги использовали ту же стратегию, которую мы рассматривали ранее в этой главе для компьютерного моделирования стай скворцов. Вместо того, чтобы программировать поведение всей бластулы, они запрограммировали единственную клетку. Затем они "клонировали" много клеток, все время наблюдая за тем, что происходило, когда эти клетки собрались вместе на компьютере. Когда я говорю, что они запрограммировали поведение единственной клетки, будет лучше сказать, что они запрограммировали математическую модель единственной клетки, собрав в этой модели некоторые известные факты об одиночной клетке, но в упрощенной форме. В частности, известно, что внутренние пространства клеток пересекаются микронитями: своего рода миниатюрными резинками, но с дополнительным свойством - способностью активно сокращаться, как сжимаются мышечные волокна. На самом деле, микронити используют тот же принцип сокращения, что и мышечные волокна. Модель Остера упростила клетку до двух измерений, для того чтобы изобразить ее на экране компьютера, и всего лишь с полудюжиной нитей, стратегически размещенных в клетке, как Вы видите на диаграмме выше. В компьютерной модели всем микронитям дали определенные количественные свойства с названиями, которые что-то означают для физиков: "коэффициент вязкостного демпфирования" и "постоянная упругой пружины". Не важно, что они в точности подразумевают: они являются теми вещами, которые физикам нравится измерять у пружин. Хотя надо полагать, что в реальной клетке многие нити были бы способны сокращаться, Остер и его коллеги упростили дело, снабдив только одну из шести нитей этой способностью. Если бы они смогли бы получить реалистичные результаты даже после того, как откинули некоторые из известных свойств клетки, по-видимому, было бы возможно добиться не менее хороших результатов с более сложной моделью, в которой эти свойства сохранялись. Вместо того, чтобы позволять одной сокращающейся нити в их модели произвольно сокращаться, они встроили свойство, распространенное в определенного рода мышечных волокнах: растянутое сверх определенной критической длины, волокно реагировало, сокращаясь до намного меньшей длины, чем его нормальная равновесная длина.
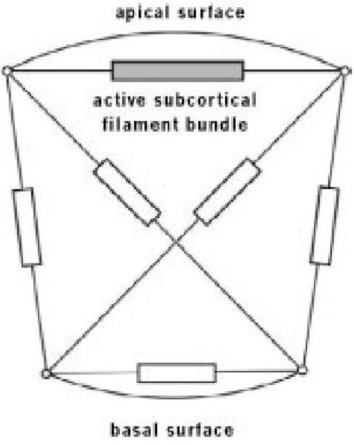
Микронити в модельной камере Остера
Итак, у нас есть своя модель единственной клетки: очень упрощенная модель, состоящая из двумерного контура, в котором натянуто шесть упругих пружин, у одной из которых есть особое свойство реагировать на приложенное извне растягивание активным сокращением. Это первый этап процесса моделирования. Во второй стадии Остер и его коллеги клонировали несколько дюжин своих моделируемых клеток и расположили их по кругу, подобно (двумерной) бластуле. Затем они взяли одну клетку и подрегулировали ее сокращающуюся нить, чтобы побудить ее к сокращению. То, что затем произошло, едва ли не слишком замечательно. Моделируемая бластула гаструлировала! Вот шесть скриншотов, показывающих, что произошло (от a до f ниже). Волна сокращений распространилась в стороны от возбужденной клетки, и шар клеток самопроизвольно вогнулся.
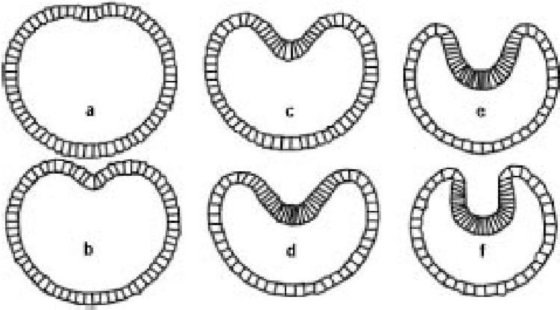
Остеровская модельная blastula gastrulating
То к чему идет - еще лучше. Остер и его коллеги поставили эксперимент на своей компьютерной модели, в котором понизили "порог срабатывания" сокращающихся нитей. Результатом была волна инвагинации, которая пошла дальше и фактически отщепила нервную трубку (скриншоты от а до h на обороте). Важно понять, чем реально является модель, такая как эта. Она - не точное представление нейруляции. Помимо того факта, что она являлся двумерной и упрощенной во многих других отношениях, шар клеток, который нейрулировал (скриншот а), не был двухслойной "гаструлой", как это должно было быть. Это была все та же бластулоподобная исходная позиция, какая была у нас в модели гаструляции выше. Это не имеет значения: не следует думать, что модели должны быть абсолютно точны во всех деталях. Модель, тем не менее, показывает, как легко воспроизвести различные аспекты поведения клеток в раннем эмбрионе. Факт, что двумерный "шар" клеток самопроизвольно отреагировал на стимул, даже при том, что модель более проста, чем реальная ситуация, делает ее более сильным свидетельством. Он убеждает нас, что эволюция различных процедур раннего эмбрионального развития не обязана быть такой уж сложной. Заметьте, что простой является модель, а не явление, которое она демонстрирует. Это является признаком хорошей научной модели.
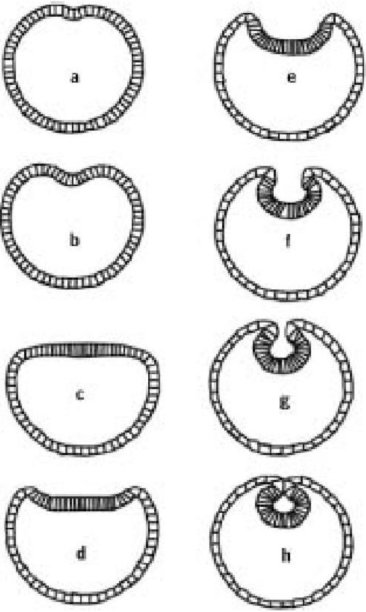
Формирование 'нервной трубки' в модели Остера
Моя цель разъяснения моделей Остера состояла в том, чтобы показать общий принцип, благодаря которому отдельные клетки могут взаимодействовать друг с другом, строя тело без какого-либо чертежа, отображающего целое тело. Подобно складыванию оригами, инвагинация и отделение в стиле [эксперимента] Остера являются лишь некоторыми из самых простых уловок для построения эмбрионов. Другие, более сложные, играют роль позже в эмбриональном развитии. Например, изобретательные эксперименты показали, что нервные клетки, когда растут от спинного или головного мозга, находят дорогу к своему конечному органу, не следуя какого-либо рода общему плану, а благодаря химическому привлечению, почти как пес вынюхивает, выискивая суку в период течки. Ранний классический эксперимент Нобелевского лауреата, эмбриолога Роджера Сперри отлично иллюстрирует этот принцип. Сперри с коллегой взяли головастика и удалили крошечный квадратик кожи со спины. Они удалили другой квадратик, такого же размера, с живота. Затем они пересадили эти два квадратика, но каждый на место другого: кожа живота была пересажена на спину, а кожа спины - на живот. Когда головастик вырос в лягушку, результат был довольно симпатичен, как часто бывает с экспериментами в эмбриологии: имелась аккуратная почтовая марка белой кожи живота посреди темной, пестрой спины, а другая аккуратная почтовая марка темной пятнистой кожи - посреди белого живота. А теперь - главное в этой истории. Обычно, если пощекотать спину лягушки щетинкой, лягушка будет чесать это место лапой, как будто отгоняя раздражающую муху. Но когда Сперри щекотал своей экспериментальной лягушке белую "почтовую марку" на спине, она чесала живот! А когда Сперри щекотал ей темную почтовую марку на животе, лягушка чесала свою спину.
В нормальном эмбриональном развитии, согласно интерпретации Сперри, произошло следующее: аксоны (длинные "провода", каждый - узкий, трубчатый выступ отдельной нервной клетки) вырастают из спинного мозга в поисках кожи живота, вынюхивая ее как собака. Другие аксоны растут из спинного мозга, вынюхивая кожу спины. И обычно это дает правильный результат: щекотание спины ощущается, как если бы оно было на спине, в то время как щекотание живота ощущается, как если бы оно было на животе. Но у экспериментальной лягушки Сперри некоторые из нервных клеток, вынюхивающих кожу живота, нашли почтовую марку кожи живота пересаженной на спину, по-видимому, потому что она правильно пахла. И наоборот. Люди, которые верят в своего рода теорию tabula rasa (чистой доски), согласно которой мы все рождены с разумом в виде с чистого листа, и заполняем его опытом, должны быть удивлены результатом Сперри. Они должны ожидать, что лягушки будут учиться на опыте разбираться с ощущениями их собственной кожи, связывая правильные чувства с правильными местами на коже. Вместо этого кажется, что каждая нервная клетка в спинном мозге помечена, скажем, как нервная клетка живота или нервная клетка спины, даже прежде, чем она вступит в контакт с соответствующей кожей. Позже она найдет свой назначенный, целевой пиксель кожи, где бы он ни был. Если бы муха проползла вдоль ее спины, лягушка Сперри, по-видимому, испытала бы иллюзию, что муха внезапно перепрыгнула со спины на живот, проползла немного дальше, а затем мгновенно перепрыгнула снова на спину.
Подобные эксперименты вынудили Сперри сформулировать свою гипотезу "хемо-афинности", согласно которой нервная система монтируется, не следуя всеобщему чертежу, а благодаря тому, что каждый отдельной аксон, ищет конечные органы, с которыми у него есть особое химическое сродство. Еще раз, у нас есть маленькие, локальные единицы, придерживающиеся локальных правил. Клетки обычно изобилуют "ярлыками", химическими значками, которые позволяют им найти своих "партнеров". И мы можем вернуться к аналогии оригами, чтобы найти другое место, где принцип маркировки оказывается полезным. В человеческом бумажном оригами не используется клей, хотя мог бы. А в оригами эмбриона, в соответствии с которым собираются тела животных, действительно использует что-то аналогичное клею. Скорее клеям, потому что их существует много, и именно здесь маркировка торжествующе получает должное признание. У клеток есть сложный ассортимент "молекул адгезии" на их поверхностях, посредством которых они липнут к другим клеткам. Это клеточное слипание играет важную роль в эмбриональном развитии во всех частях тела. Тем не менее, есть существенное отличие от клеев, с которыми мы знакомы. Для нас клей является всего лишь клеем. Некоторые клеи сильнее других, некоторый из клеев схватываются быстрее других, некоторые клеи больше подходят, скажем, для дерева, в то время как другие лучше работают с металлами или пластмассами. Но это, пожалуй, и все относительно разнообразия среди клеев.
Молекулы клеточной адгезии работают намного более тонко. Можно сказать, они более разборчивы. В отличие от наших искусственных клеев, которые будут клеить большинство поверхностей, молекулы клеточной адгезии связываются только с другими особыми молекулами клеточной адгезии в точности правильной разновидности. Один класс адгезивных молекул у позвоночных, кадгерины, имеется в приблизительно восьмидесяти известных в настоящее время специфичных разновидностях. За некоторыми исключениями, каждый из этих приблизительно восьмидесяти кадгеринов свяжется только со своей собственной разновидностью. Забудьте на минутку клей: лучшей аналогией могла бы быть игра на детском празднике, где каждому ребенку назначают животное, и все они должны, кружа по комнате, производить звуки, как отведенные им животные. Каждый ребенок знает, что только одному из других детей назначили то же животное, что и ему, и он должен найти своего партнера, слушая какофонию, имитирующую фермерский двор. Подобно работают и кадгерины. Возможно, как и я, Вы смутно можете представить, как толковое добавление конкретных кагдеринов на поверхности клеток в стратегически важных местах могло бы улучшить и усложнить принципы самосборки оригами эмбриона. Заметьте, снова же, что это подразумевает не какой бы то ни было всеобщий план, а скорее разрозненную коллекцию локальных правил.
ФЕРМЕНТЫ
Видя, как целые слои клеток играют в игру оригами при формировании эмбриона, давайте теперь погрузимся в отдельную клетку, где мы обнаружим тот же принцип самосворачивания и самосгибания, но в намного меньшем масштабе, масштабе отдельной молекулы белка. Белки крайне важны по причинам, разъяснению которых мне следует уделить время, начиная с дразнящего рассуждения, превозносящего уникальную важность белков. Я люблю размышлять о том, насколько причудливо отличной, следует ожидать, будет жизнь в другом месте вселенной, но есть одна или две вещи, которые, я подозреваю, универсальны везде, где бы ни была обнаружена жизнь. Все живое окажется эволюционировавшим благодаря процессу, имеющему отношение к дарвиновскому естественному отбору генов. И она будет сильно опираться на белки - или молекулы, которые, как белки, способны сворачиваться в огромное разнообразие форм. Молекулы белка - виртуозы автооригамного искусства, в масштабе, намного меньшем, чем масштаб слоев клеток, с которыми мы пока что имели дело. Молекулы белка - поразительный образец того, что может быть достигнуто при следовании локальным правилам в локальном масштабе.
Белки - цепочки меньших молекул, называемых аминокислотами, и эти цепочки, подобно слоям клеток, которые мы рассматривали, также сворачиваются весьма строго заданным способом, но в намного меньшем масштабе. В белках, встречающихся в природе (это - один факт, который, по-видимому, будет другим в инопланетных мирах) есть только двадцать разновидностей аминокислот, и все белки являются цепочками, собранными лишь из этого ассортимента двадцати, выделенного из намного большего набора возможных аминокислот. Теперь - к автооригами. Белковые молекулы, просто следуя законам химии и термодинамики, спонтанно и автоматически скручиваются в трехмерные конфигурации точной формы - я почти сказал "узлы", но, в отличие от миксин (если я могу необоснованно сообщить не относящийся к делу, но занимательный факт), белки не завязываются буквально в узлы. Трехмерная структура, в которую загибается и скручивается цепочка белка, является "третичной структурой", с которой мы кратко познакомились, рассматривая самосборку вирусов. Любая конкретная последовательность аминокислот диктует специфическую схему сворачивания. Последовательность аминокислот, в свою очередь определяемая последовательностью букв в генетическом коде, определяет форму третичной структуры. Форма третичной структуры, в свою очередь, имеет чрезвычайно важные химические последствия.
Автооригами, в соответствии с которым сворачиваются и закручиваются цепочки белка, управляются при помощи законов химической притягательности и законов, определяющих углы, под которыми атомы связаны друг с другом. Вообразите ожерелье из магнитов необычной формы. Это ожерелье не висело бы грациозно прогибающейся линией вокруг грациозной шеи. Оно приняло бы какую-то другую форму, став запутанным, поскольку магниты удерживали бы друг друга и нашли бы друг у друга уголки и щели в различных местах вдоль цепочки. В отличие от случая с цепочкой белка, точная форма переплетения не была бы предсказуемой, потому что любой магнит притягивает любой другой. Но она иллюстрирует, как цепочки аминокислот могут спонтанно формировать сложную узловатую структуру, которая может быть не похожа на цепь или ожерелье.
Подробности того, как законы химии определяют третичную структуру белка, полностью еще не поняты: химики еще не могут установить во всех случаях, как свернется данная последовательность аминокислот. Однако есть хорошие свидетельства, что третичная структура в принципе выводима из последовательности аминокислот. Нет ничего таинственного во фразе "в принципе". Никто не может предсказать, как упадет игральная кость, но все мы верим, что это всецело определено точными деталями того, как она брошена, плюс некоторыми дополнительными фактами о сопротивлении воздуха и так далее. Проверенный факт, что определенная последовательность аминокислот всегда сворачивается в определенную форму или одну из дискретного набора альтернативных форм (см. длинную сноску напротив). И - важный вопрос для эволюции - сама последовательность аминокислот полностью определена, через применение правил генетического кода, последовательностью (триплетами) "букв" в гене. Даже при том, что смертным химикам нелегко предсказать, какое изменение в форме белка будет следствием конкретной генетической мутации, тем не менее, существует факт, что, как только мутация произошла, итоговое изменение формы белка будет в принципе предсказуемо. Один и тот же мутантный ген надежно произведет одну и ту же измененную форму белка (или дискретное меню альтернативных форм). И это -все, что имеет значение для естественного отбора. Естественному отбору не нужно понимать, почему генетическое изменение имеет определенное последствие. Достаточно, что оно есть. Если это последствие влияет на выживание, то сам измененный ген погибнет или уцелеет при конкуренции за доминирование в генофонде, независимо от того, понимаем ли мы точный путь, которым ген влияет на белок.
Учитывая, что форма белков очень разнообразна, и учитывая, что она определена генами, почему она так в высшей степени важна? Частично, потому что некоторые белки играют в теле непосредственно структурную роль. Фибриллярные белки, такие как коллаген, объединяются в крепкие веревки, которые мы называем связками и сухожилиями. Но большинство белков не является фибриллярными. Вместо этого они сворачиваются в свою характерную шаровидную форму с едва уловимыми впадинами, и эта форма определяет характерную роль белка как фермента, то есть катализатора.
Катализатор - химическое вещество, которое ускоряет, в миллиарды или даже триллионы раз, химическую реакцию между другими веществами, в то время как сам катализатор выходит из процесса, невредимым и готовым катализировать снова. Ферменты, которые являются белками-катализаторами - чемпионы среди катализаторов благодаря своей специфичности: они очень разборчивы в отношении того, какие химические реакции они ускоряют. Или, пожалуй, можно сказать: химические реакции в живых клетках очень разборчивы в отношении ускоряющих их ферментов. Многие реакции в клеточной химии настолько медленны, что без подходящего фермента практически не происходят вообще. Но с нужным ферментом они протекают очень быстро и могут производить продукты в большом количестве.
Вот как мне нравится это выражать. В химической лаборатории на полках есть сотни бутылок и банок, каждая содержит различные чистые вещества: соединения и элементы, растворы и порошки. Химик, желающий произвести определенную химическую реакцию, выбирает две или три бутылки, берет порцию из каждой, смешивает их в пробирке или колбе, возможно, нагревает, и происходит реакция. Другие химические реакции, которые могли бы произойти в лаборатории, не происходят, потому что стеклянные стенки бутылок и колб препятствуют встрече компонентов. Если Вы хотите другую химическую реакцию, Вы смешиваете другие компоненты в другой колбе. Повсюду есть стеклянные барьеры, сохраняющие вещества чистыми, отдельными друг от друга в бутылках или колбах, и содержащие комбинации реагентов отдельно друг от друга в пробирках, или колбах, или мензурках.
Живая клетка также является большой химической лабораторией, и в ней есть такой же большой запас химикатов. Но они не хранятся в отдельных бутылках и банках на полках. Они все перемешаны. Это как если бы вандал, химический владыка хаоса, вошел бы в лабораторию, схватил все бутылки на всех полках и опрокинул их с разнузданной импульсивностью в один большой котел. Ужасно? Что ж, было бы ужасно, если бы все они прореагировали вместе, во всех возможных комбинациях. Но они не реагируют. Или если они и реагируют, то скорость, с которой они это делают, настолько мала, что с тем же успехом они могли бы не реагировать вообще. Кроме как - и в этом вся суть - если присутствует фермент. Нет никакой надобности в стеклянных бутылках и колбах, чтобы содержать вещества отдельно, потому что в любом случае они не будут реагировать вместе как попало - пока не появится подходящий фермент. Эквивалентом хранения химикатов в закупоренных бутылках, прежде чем Вы захотите смешать отдельную пару, скажем A и B, является смешивание всех сотен веществ в большом ведьмином вареве, но с добавкой только фермента, нужного чтобы катализировать реакцию между A и B, и никакую другую комбинацию. На самом деле метафора склонных к беспорядочности опрокинутых бутылок заходит слишком далеко. Клетки в действительности содержат инфраструктуру мембран, между которыми, и в пределах которых, и происходят химические реакции. До некоторой степени эти мембраны играют роль стеклянных перегородок между пробирками и колбами.
Суть этого раздела этой главы в том, что "правильный фермент" достигает своей "правильности" в значительной степени вследствие своей физической формы (и это важно, потому что физическая форма определяется генами, и это - гены, чьим вариациям, в конечном счете, благоприятствует или не благоприятствует естественный отбор). Молекулы в большом количестве дрейфуют, вертятся и кружат в бульоне, омывающем внутренности клетки. Молекула вещества A могла бы быть счастлива прореагировать с молекулой вещества B, но только если им доведется столкнуться, будучи повернутыми в строго правильном направлении друг относительно друга. Важно то, что это редко случается - если в процесс не вступает подходящий фермент. Точная форма фермента, форма, в которую он свернулся как магнитное ожерелье, оставляет его изрытым впадинами и вмятинами, у каждой из которых есть своя точная форма. У каждого фермента есть так называемый "активный центр", который обычно представляет собой специфическую впадину или карман, чья форма и химические свойства придают ферменту его специфику. Слово "впадина" недостаточно передает особенность, точность, этого механизма. Возможно, лучшее сравнение с электрической розеткой. В том, что мой друг зоолог Джон Кребс назвал "великим штепсельным заговором", различные страны во всем мире, что раздражает, приняли разные произвольные соглашения для штепселей и розеток. Британские штепсели не будут подходить к американским или французским розеткам, и так далее. Активные центры на поверхности молекул белка - розетки, которым будут отвечать только определенные молекулы. Но если великий штепсельный заговор ведет только к полудюжине отдельных форм во всем мире (вполне достаточно, чтобы создать постоянные неудобства путешественнику), различные виды розеток, представляемых ферментами, намного более многочисленны.
Представьте себе конкретный фермент, который катализирует химическую комбинацию двух молекул, P и Q, создавая соединение PQ. Одна половина активного центра "розетки" как раз подходит, чтобы в нем устроилась молекула типа P, как часть пазла. Другая половина той же розетки столь же точно приспособлена, чтобы в нее вставлялась молекула Q - обращенная в точности в правильном направлении, чтобы химически соединиться с молекулой P, уже находящейся там. Находясь в одной впадине, твердо удерживаемые как раз под нужным углом друг к другу сводящей молекулой фермента, P и Q объединяются. Новое соединение, PQ теперь отделяется в бульон, оставляя активную впадину в молекуле фермента готовой примирять другую P и другую Q. Клетка может быть заполнена роем идентичных молекул фермента, продолжающих работать как роботы на автомобильном заводе, производя в большом количестве PQ в клеточном аналоге промышленных количеств. Поместите другой фермент в одну и ту же клетку, и он произведет в большом количестве другой продукт, возможно PR, или QS или YZ. Конечные продукты различны, даже при том, что доступное сырье -одно и то же. Другие типы ферментов заняты не созданием новых соединений, а расщеплением старых. Некоторые из этих ферментов вовлечены в переваривание пищи, и они используются также в "биологических" стиральных порошках. Но, так как эта глава о строительстве эмбрионов, мы здесь рассматриваем главным образом строительные ферменты, которые посредничают в синтезе новых химических соединений. Один такой процесс показан в действии на цветной странице 12.
Проблема могла прийти Вам в голову. Все это очень хорошо, говорить о пазле впадин и розеток, высокоспецифичных активных центрах, способных ускорить конкретную химическую реакцию в триллионы раз. Но разве это не звучит слишком хорошо, чтобы быть правдой? Как молекулы фермента как раз нужной формы эволюционировали из менее совершенных начал? Какова вероятность, что у розетки, сформированной случайным образом, будет как раз нужная форма и как раз нужные химические свойства, чтобы обеспечить соединение между двумя молекулами, P и Q, ловко устраивая их столкновение под точно правильным углом? Не очень большая, если Вы представите себе "законченный пазл" - или, на самом деле, если Вы представите себе "великий штепсельный заговор". Вместо этого Вы должны представить себе "плавный градиент усовершенствования". Поскольку нередко, когда мы сталкиваемся с загадкой, как сложные и невероятные вещи могут явиться результатом эволюции, ошибочно предполагать, что итоговое совершенство, которое мы видим сегодня, было всегда. Полностью сформированные, сильно эволюционировавшие молекулы фермента обеспечивают увеличение в триллион раз скорости реакций, которые они катализируют, и они делают это за счет свей красивой, как раз подходящей формы. Но Вам не нужно увеличение скорости в триллион раз, чтобы вам благоприятствовал естественный отбор. Вполне устроит и в миллион раз! Подойдет и тысяча раз. И даже в десять или в два раза было бы достаточно естественному отбору для надлежащего закрепления. Есть плавный градиент усовершенствования эффективности фермента на всем протяжении от почти полного отсутствия впадины, через впадину грубой формы, до розетки совершенно правильной формы и химических деталей. "Градиент" означает, что каждый шаг является заметным усовершенствованием, каким бы небольшим он ни был, относительно предыдущего. И "заметное" для естественного отбора может означать усовершенствование, меньшее, чем тот минимум, который был бы необходим нам, чтобы его заметить.
Итак, Вы видите, как это работает. Изящно! Клетка -переналаживаемая химическая фабрика, способная выплевывать в массовых количествах широкое разнообразие различных веществ, выбранных под влиянием того, какой присутствует фермент. И как делается этот выбор? В соответствии с тем, какой ген включен. Так же как клетка является чаном, заполненным большим количеством химикатов, только меньшая часть которых реагирует друг с другом, так же и каждое клеточное ядро содержит весь геном, но только меньшая часть генов включена. Когда ген включен, скажем, в клетке поджелудочной железы, его последовательность кодовых букв непосредственно определяет последовательность аминокислот в белке; а последовательность аминокислот определяет (помните образ магнитного ожерелья?) форму, в которую сворачивается белок; а форма, в которую сворачивается белок, определяет точную форму впадин, которые соединяют вещества, дрейфующие вокруг в клетке. Каждая клетка, за очень немногими исключениями, такими как красные кровяные тельца, не имеющие ядра, содержит гены для того, чтобы производить все ферменты. Но в любой клетке только несколько генов будут включены в любое конкретное время. Скажем, в клетках щитовидной железы гены, которые продуцируют ферменты, подходящие для катализа производства гормонов щитовидной железы, включены. И соответственно для всех различных видов клеток. Наконец, химические реакции, протекающие в клетке, определяют форму клетки, и особенность ее поведения, и то, как она участвует в взаимодействиях в стиле оригами с другими клетками. Таким образом, всем течением эмбрионального развития управляют, через запутанную последовательность событий, гены. Это гены, которые определяют последовательности аминокислот, которые определяют третичные структуры белков, которые определяют розеткоподобные формы активных центров, которые определяют химию клетки, которые определяют "подобное скворцам" поведение клетки в эмбриональном развитии. Так различия в генах могут служить причиной, в начальном конце сложной цепи событий, различий в том, как развивается эмбрион и, следовательно, различий в форме и поведении взрослых. Выживание и репродуктивный успех этих взрослых в таком случае действуют обратной связью в выживании в генофонде генов, определивших разницу между успехом и неудачей. И это - естественный отбор.

Клеточная родословная Caenorhabditis elegans
Эмбриология кажется сложной - является сложной - но легко понять важную суть, состоящую в том, что мы на всем пути имеем дело с локальными процессами самосборки. Отдельный вопрос, учитывая, что (почти) все клетки содержат все гены, как определяется, какие гены включаются в каждом различном виде клеток. Теперь я должен кратко рассмотреть этот вопрос.
ЗАТЕМ ПОСТАРАЮТСЯ ЧЕРВИ
Включен ли данный ген в данной клетке в данное время, часто определяется посредством каскада других генов, названных генами переключателями или регуляторными генами, под влиянием химического окружения клетки. Клетки щитовидной железы весьма отличаются от мышечных клеток и т. д., даже при том, что их гены -одни и те же. Все это очень хорошо, Вы можете сказать, если только развитие эмбриона идет полным ходом, и различные виды тканей, такие как щитовидная железа и мышцы, уже существуют. Но каждый эмбрион начинает свою жизнь как одна клетка. Клетки щитовидной железы и клетки мышц, клетки печени и клетки костей, клетки поджелудочной железы и клетки кожи, все происходят от одной оплодотворенной яйцеклетки через ветвящееся генеалогическое дерево. Это генеалогическое дерево клеток, берущее начало не далее чем в момент оплодотворения, не имеет ничего общего с эволюционным деревом, берущим начало миллионы лет назад, которое продолжает появляться в других главах. Позвольте мне показать Вам, например, полное генеалогическое дерево всех 558 клеток недавно рожденной личинки червя нематоды, Caenorhabditis elegans (ниже: пожалуйста, обратите особое внимание на каждую деталь этой диаграммы). Между прочим, я не знаю, что совершил этот крошечный червь, чтобы заработать свое видовое название elegans, но я могу подумать о серьезном основании, почему он мог его заслужить ретроспективно. Я знаю, что не всем моим читателям нравятся мои отступления, но исследование, которое было сделано на Caenorhabditis elegans является таким звучным триумфом науки, что Вам меня не остановить.
Caenorhabditis elegans был выбран в 1960-ых как идеальное экспериментальное животное потрясающе блистательным южноафриканским биологом Сиднеем Бреннером. Он недавно закончил свою работу с Фрэнсисом Криком и другими в Кембридже по расшифровке генетического кода и искал, какую бы новую большую проблему решить. Его вдохновенный выбор и его собственное исследования в области генетики и нейроанатомии привели к международному сообществу исследователей Caenorhabditis, которое разрослось до тысяч. Было бы некоторым преувеличением сказать, что мы теперь знаем все о Caenorhabditis elegans! Мы знаем весь его геном. Мы знаем точно, где каждая из его 558 клеток (в личинке; 959 во взрослой гермафродитной форме, не считая репродуктивных клеток), находится в теле, и мы знаем точную "семейную историю" каждой из этих клеток на протяжении эмбрионального развития. Мы знаем о большом количестве мутантных генов, которые производят неправильных червей, и мы знаем точно, где мутация действует в теле и точную клеточную историю того, как аномалия развивается. Это небольшое животное известно от начала до конца, известно наизнанку, известно от головы до хвоста и всех промежуточных станций, известно вдоль и поперек ("O замечательный день!"). Бреннер был с запозданием удостоен Нобелевской премии по физиологии в 2002 году, а родственный вид назвали в его честь, Caenorhabditis brenneri. Его постоянная рубрика в журнале "Current Biology", подписываемая "Дядя Сид", является образцом интеллектуального и непочтительного научного остроумия - столь же изящного, как международные усилия в исследовании C. elegans, которые он вдохновил. Я бы хотел, чтобы молекулярные биологи поговорили бы с некоторыми зоологами (как сам Бреннер) и научились бы не называть Caenorhabditis просто нематода или даже просто червь, как если бы не было никаких других.
Конечно, Вы не можете прочитать названия типов клеток у основания диаграммы (потребовалось бы семь страниц, чтобы распечатать все это разборчиво), но они перечисляют такие вещи как "глотка", "кишечная мышца", "мышца тела", "мышца сфинктера", "кольцевой ганглий", "люмбальный ганглий". Клетки всех этих типов являются буквально кузенами друг друга: кузенами по своему происхождению за время жизни отдельного червя. Например, я смотрю на особую мышечную клетку тела, названную MSpappppa, которая является родной сестрой другой мышечной клетки тела, двоюродной сестрой еще двух мышечных клеток тела, племянницей еще двух мышечных клеток тела, троюродной сестрой шести клеток глотки, четвероюродной сестрой семнадцати клеток глотки... и так далее. Не удивительно ли, что мы можем фактически использовать такие слова как "троюродная племянница", с предельной точностью и уверенностью, чтобы указывать на проименованные и однозначно идентифицируемые клетки тела животного? Число "поколений" клеток, отделяющее ткани от первоначального яйца, не особенно велико. В конце концов, в этом теле всего лишь 558 клеток, и Вы можете теоретически создать 1 024 (2 в 10 степени) за десять поколений клеточного деления. Число поколений для клеток человека было бы намного большим. Однако теоретически можно было бы сделать подобное генеалогическое дерево для каждой из триллиона с лишним Ваших клеток (в отличие от 558 клеток личинки самки C.elegans), прослеживая родословную каждой назад к одной оплодотворенной яйцеклетке. У млекопитающих, однако, невозможно идентифицировать конкретные, однозначно проименованные клетки. У нас больше имеют место статистические популяции клеток, чьи детали различны у разных людей.
Я надеюсь, мое эйфорическое отступление к элегантности исследования Caenorhabditis не слишком нас отвлекло от проводимого мною рассуждения о том, как меняются типы клеток по своей форме и характеру по мере их ответвления друг от друга в эмбриональном генеалогическом дереве. В точке ветвления между клоном, которому предстоит стать клетками глотки, и " клоном-кузеном", которому предстоит стать клетками кольцевого ганглия, должно быть нечто, что отличало бы их, иначе как бы они сумели включить различные гены? Ответ в том, что когда последний общий предок двух клонов разделился, две половины клетки перед делением были различными. Поэтому, когда клетка разделилась, две дочерних клетки, хотя идентичные в своих генах (каждая дочерняя клетка получает полный комплект генов), не были идентичны в окружающих химикатах. И это означает, что были включены не одни и те же гены, что изменило судьбу их потомков. Тот же принцип применяется сплошь в эмбриологии, включая самое ее начало.
Ключом к дифференциации у всех животных является асимметричное деление клетки.
Сэр Джон Салстон и его коллеги проследили каждую из клеток в теле червя назад к одной и только одной из шести клеток-основательниц -мы могли бы даже назвать их "клетками-матриархами" - названных AB, MS, E, D, C и P4. Называя клетки, они использовали аккуратные обозначения, которые кратко подытоживают историю каждой. Название каждой клетки начинается с названия одной из тех шести клеток-основательниц, той, от которой она происходит. Дальнейшее ее название является последовательностью букв, начальных букв адреса отдела клеток, давшего ей начало: передний, задний, спинной, брюшной, левый, правый. Например, Ca и Cp являются двумя дочерьми матриарха C, передней [anterior] и задней [posterior] дочерьми соответственно. Заметьте, что у каждой клетки есть не больше двух дочерей (из которых одна может умереть). Теперь я смотрю на конкретную мышечную клетку тела, название которой, Cappppv, кратко раскрывает ее историю: у C была передняя дочь, у которой была задняя дочь, у которой была задняя дочь, у которой была задняя дочь, у которой была задняя дочь, у которой была брюшная [ventral] дочь, которая является рассматриваемой мышечной клеткой тела. Каждая клетка в теле обозначена сопоставимой последовательностью букв, возглавляемой одной из шести клеток-основательниц. Возьмем другой пример, ABprpapppap является нервной клеткой, которая находится в брюшном нервном стволе, идущем вдоль червя. Стоит ли говорить, что нет необходимости вдаваться в детали. Красивый момент в том, что у каждой отдельной клетки тела есть такое название, которое полностью описывает ее историю во время эмбриологенеза. Каждое из десяти делений клетки, давших начало ABprpapppap и любой другой клетке, было асимметричным делением с возможностью различных генов быть включенными в каждой из двух дочерних клеток. И у всех животных это является принципом, благодаря которому дифференцируются ткани, даже при том, что все их клетки содержат одни и те же гены. У большинства животных, конечно, гораздо больше клеток, чем 558 клеток Caenorhabditis, и их эмбриональное развитие в большинстве случаев менее жестко детерминировано. В частности, как мне любезно напомнил сэр Джон Салстон, и как я уже вкратце упомянул, у млекопитающего "генеалогические деревья" наших клеток различны для каждой особи, тогда как в Caenorhabditis они почти идентичны (кроме особей-мутантов). Однако принцип остается одним и тем же. У любого животного клетки отличаются друг от друга в различных частях тела, даже хотя они и генетически идентичны, из-за их истории асимметричного клеточного деления во время короткого курса эмбрионального развития.
Послушаем же окончание всей этой истории. Нет никакого всеобщего плана развития, никакого чертежа, никакого архитектурного плана, никакого архитектора. Развитие эмбриона и, в конечном счете, взрослого достигается благодаря локальным правилам, применяемым клетками, взаимодействующими с другими клетками на локальной основе. То, что происходит внутри клеток, подобным же образом управляется локальными правилами, которые касаются молекул, особенно белковых молекул, внутри клеток и в клеточных мембранах, взаимодействующих с другими такими молекулами. Снова же, все правила являются локальными, локальными, локальными. Никто, читая последовательность букв в ДНК оплодотворенного яйца, не смог бы предсказать форму животного, в которую оно собирается вырасти. Единственный способ узнать это - вырастить яйцо естественным способом и увидеть то, во что оно превратится. Никакой электронный компьютер не смог бы решить это, если он не был непосредственно запрограммирован моделировать естественный биологический процесс, и в этом случае Вы могли бы с тем же успехом обойтись без электронной версии и использовать развивающийся эмбрион как его собственный компьютер. Этот способ образовывать большие и сложные структуры лишь выполняя локальные правила, очень отличается от метода делать вещи по чертежу. Если бы ДНК была некоторым линеаризованным чертежом, то было бы относительно тривиальной задачей запрограммировать компьютер, чтобы прочитать буквы и начертить животное. Но было бы совсем нелегко - в действительности могло бы быть невозможно - для животного эволюционировать изначально.
И теперь, чтобы эта глава об эмбрионах не заканчивалась как простое отступление в книге по эволюции, я должен вернуться к подлинной дилемме спросившей Холдейна леди. Учитывая, что гены управляют процессами эмбрионального развития, а не взрослой формой; учитывая, что естественный отбор - подобно Богу - не строит крошечные крылья, а эмбриология строит; как естественный отбор воздействует на животных, формируя их тела и их поведение? Другими словами, как естественный отбор воздействует на эмбрионы, перестраивая их так, что они становятся все более искусными в построении успешных тел, с крыльями или плавниками, листьями или броневой защитой, жалами или щупальцами, или чем бы то ни было, что требуется для выживания?
Естественный отбор - различное выживание успешных генов по сравнению с альтернативными, менее успешными генами в генофондах. Естественный отбор не выбирает гены непосредственно. Вместо этого он выбирает их полномочных представителей, тела особей; и эти особи выбираются - явно и автоматически, и без умышленного вмешательства - тем, выживают ли они, чтобы воспроизвести копии тех самых генов. Выживание гена тесно переплетается с выживанием тел, которые он помогает строить, потому что он ездит в этих телах и умирает с ними. Любой конкретный ген может рассчитывать оказаться, в форме своих копий, едущим в большом количестве тел одновременно в популяции современников, по мере того как одно поколение сменяет другое. Статистически, поэтому, ген, склонный в среднем оказывать положительный эффект на перспективы выживания тел, в которых он оказывается, будет иметь тенденцию увеличивать свою частоту в генофонде. Так, в среднем, гены, с которыми мы встретим в генофонде, будут как правило теми генами, которые хорошо строят тела.
Эта глава была о процедурах, посредством которых гены строят тела. Собеседница Холдейна нашла неправдоподобным, что естественный отбор мог составить, скажем, за миллиард лет генетический рецепт для ее построения. Я нахожу это правдоподобным, хотя, конечно, ни я, ни кто-либо еще не можем сообщить Вам детали того, как это случилось. Причина, вероятно, именно в том, что это все сделано по локальным правилам. В любом акте естественного отбора мутация, которая была отобрана, имела - в большом количестве клеток и в большом количестве особей параллельно - очень простое влияние на форму, в которую спонтанно свертывается цепочка белка. Та, в свою очередь, через каталитическое действие, ускорила, скажем, конкретную химическую реакцию во всех клетках, в которых включен этот ген. Эти, возможно, изменяет скорость роста эмбрионального зачатка челюсти. А это имеет важное влияние на форму всего лица, возможно, сокращая морду и придавая ей более человеческий и менее обезьяноподобный профиль. Теперь давления естественного отбора, благоприятствующие или неблагоприятствующие гену, могут быть сколь угодно сложными. Они могут включать половой отбор, возможно, эстетический отбор высокого порядка потенциальными половыми партнерами. Или изменение в форме челюсти могло бы оказать тонкий эффект на способность животного раскалывать орехи или его способность бороться с конкурентами. Некоторая чрезвычайно сложная комбинация давлений отбора, противостоя друг другу и приходя к компромиссу с приводящей в замешательство сложностью, может влиять на статистический успех этого конкретного гена, по мере того, как он размножается в генофонде. Но ген об этом ничего не знает. Все, что он делает, в различных телах и в сменяющих друг друга поколениях, это перестраивает тщательно профилированную впадину в молекуле белка. Остаток истории следует автоматически, в ветвящихся каскадах локальных последствий, из которых, в конечном счете, появляется целое тело.
Еще более сложной, чем давления отбора в экологической, половой и социальной среде животных, является фантасмагорическая сеть влияний, которая распространяется внутри и среди развивающихся клеток: влияний генов на белки, генов на гены, белков на экспрессию генов, белков на белки; мембраны, химические градиенты, физические и химические направляющие в эмбрионах, гормоны и другие посредники действия на расстоянии, помечающие клетки, ищут другие, с идентичными или комплементарными метками. Никто не понимает картину в целом, и никто не должен понимать ее, чтобы признать изящное правдоподобие естественного отбора. Естественный отбор благоприятствует выживанию в генофонде генетических мутаций, ответственных за создание ключевых изменений в эмбрионах. Целая картина возникает как следствие сотен тысяч маленьких, локальных взаимодействий, каждое в принципе постижимо (хотя разгадать на практике может быть слишком трудно или отнимать слишком много времени) для любого, кто с достаточным терпением их исследует. На практике целое может быть непонятным и таинственным, но нет никакой тайны в принципе, ни в самой эмбриологии, ни в эволюционной истории, в результате которой регуляторные гены достигли выдающегося положения в генофонде. Сложность накапливалась постепенно в течение эволюционного времени: каждый шаг лишь чуточку отличался от предыдущего, и каждый шаг был достигнут благодаря маленьким, едва уловимым изменениям в действующих локальных правилах. Когда у Вас есть достаточное число маленьких единиц - клеток, молекул белка, мембран - каждая на своем собственном уровне повинуется локальным правилам и влияет на другие - тогда конечные последствия впечатляющи. Если гены выживают или н терпят неудачу в выживании в результате их влияния на такие локальные единицы и их поведение, то естественный отбор успешных генов - и появление их успешных продуктов - последует неизбежно. Задавшая вопрос Холдейну была неправа. В принципе не трудно сделать что-то вроде нее.
И, как сказал Холдейн, на это требуется только девять месяцев...

