Книга: Самое грандиозное шоу на Земле: доказательства эволюции
Назад: ГЛАВА 9 КОВЧЕГ КОНТИНЕНТОВ
Дальше: ГЛАВА 11 ИСТОРИЯ, НАПИСАННАЯ ПОВСЮДУ В НАС
ГЛАВА 10 ДЕРЕВО РОДСТВА
КОСТЬ К КОСТИ
Что за шедевр - скелет млекопитающего. Я хочу сказать не то, что он красив сам по себе, хотя и думаю так. Я имею в виду факт, что мы вообще можем говорить о скелете млекопитающего как таковом: факт, что столь сложно взаимосвязанная вещь настолько чудесно различна между разными видами млекопитающих во всех своих деталях и в то же время настолько очевидно одна и та же у всех млекопитающих. Наш собственный скелет достаточно нам знаком, чтобы не нужно было картинки, но посмотрите на скелет летучей мыши. Не восхитительно ли, что каждая кость имеет свой узнаваемый аналог в человеческом скелете? Узнаваемый в силу порядка соединения друг с другом. Только пропорции различаются. Руки летучей мыши сильно увеличены (относительно ее размера, конечно), но никто не упустит соответствия между нашими пальцами и длинными костями в крыльях. Человеческая рука и рука летучей мыши очевидно - ни один здравомыслящий человек не станет отрицать это - две версии одной и той же вещи. Технический термин этого вида идентичности "гомология". Машущая рука летучей мыши и наша хватательная рука гомологичны. Руки нашего общего предка -и весь его остальной скелет - были взяты и вытянуты или сдавлены, часть за частью в разных направлениях и в разной степени, среди разных линий потомков.
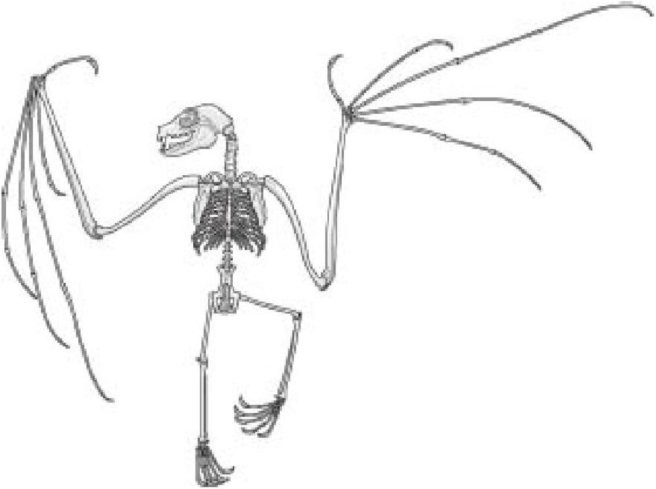
Скелет летучей мыши
То же относится - хотя, снова же, в других пропорциях - к крыльям птеродактиля (не млекопитающего, но принцип сохраняется, что еще более впечатляет). Мембрана крыла птеродактиля в основном держится на одном пальце, том что мы назвали бы мизинцем. Я признаюсь, что ощущаю невроз, навеянным гомологией, что такой вес поддерживается мизинцем, поскольку у человека он кажется столь хрупким. Глупо конечно, потому как у птеродактиля мизинец, не только не мал, но имел длину большей части туловища, и, должно быть, казался таким же сильным и крепким, как наши руки для нас. И снова, он иллюстрирует положение, которое я излагаю. Мизинец модифицирован, чтобы нести мембрану крыла. Все детали стали другими, но он по-прежнему узнаваем в силу своего пространственного расположения относительно других костей скелета. Эта длинная, крепкая, поддерживающая крыло опора гомологична нашему мизинцу. Слово "мизинец" для птеродактиля значит " весьма мощная опора".
Вдобавок к настоящим летунам - птицам, летучим мышам, птерозаврам и насекомым - многие другие животные планируют: привычка, способная поведать нам нечто о происхождении настоящего полета. У них есть мембраны для планирования, которые требуют скелетной опоры; но это не обязательно должны быть кости пальцев, как у летучих мышей и птерозавров. Летучие белки (две независимые группы грызунов), и сумчатые летяги (австралийские сумчатые, выглядящие почти в точности как летучие белки, но не близко родственные им) натягивают кожную мембрану между руками и ногами. Отдельным пальцам не требуется нести большой нагрузки и они не увеличены. Я с моим мизиничным неврозом был бы более счастлив быть летучей белкой, чем птеродактилем, так как использовать целые руки и целые ноги для несения тяжелой нагрузки ощущается "правильным".
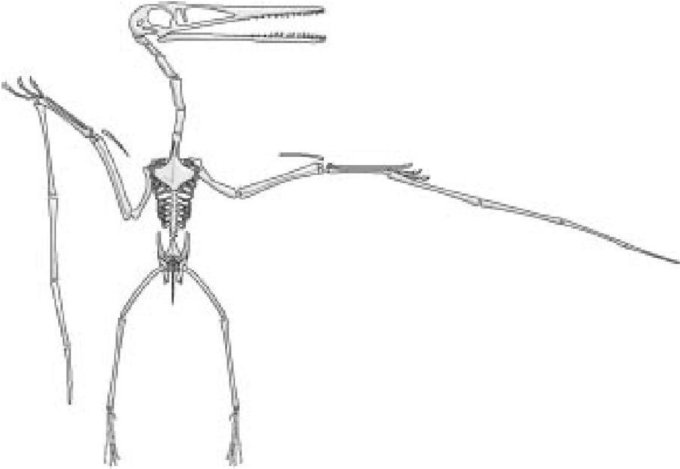
Скелет птеродактиля
На обороте представлен скелет так называемого летучего дракона, другого элегантного планера леса. Вы можете сразу увидеть, что его ребра, а не пальцы или руки и ноги модифицировались для несения "крыльев" - летательной мембраны. И еще раз, похожесть скелета в целом на скелет другого позвоночного полностью очевидна. Можно пройти по всем костям одной за другой и идентифицировать, какая кость соответствует какой в человеческом, мышином или птерозавровом скелете.
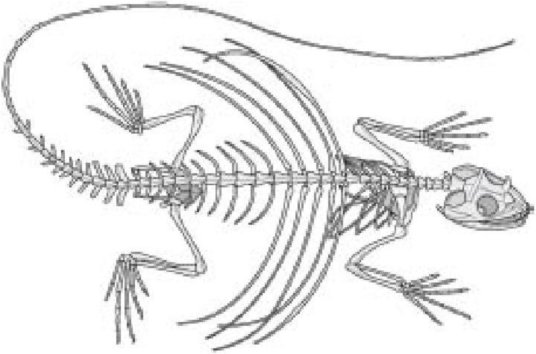
Скелет «летающей ящерицы»
Так называемые, "летучие лемуры" [шерстокрылы] лесов юговосточной Азии напоминают белок-летяг и сумчатых летяг всем, кроме того, что хвост, также как руки и ноги, включен в структуру, поддерживающую летательную мембрану. Не вызывает у меня ощущения правильности, поскольку я не могу представить, что такое иметь хвост вообще, хотя люди, как и другие бесхвостые человекообразные обезьяны, имеют рудиментарный хвост, копчик, скрытый под кожей. Для нас, почти бесхвостых обезьян, трудно себе представить, каково это быть паукообразной обезьяной, чей хвост преобладает над всем позвоночником. На рисунке на цветном листе 26 можно видеть, насколько он длиннее, даже чем длинные сами по себе руки и ноги. Как и среди многих других обезьян Нового Света (на самом деле, как и среди всех млекопитающих Нового Света вообще, что представляется курьезом, трудным для объяснения), хвост паукообразной обезьяны "хватательный", что значит, что он модифицирован для хватания, и почти кажется, что он кончается дополнительной рукой, хотя он не гомологичен настоящей руке и не имеет пальцев. На самом деле, хвост паукообразной обезьяны выглядит очень похожим на дополнительную ногу или руку.
Мне, вероятно, не нужно разъяснять идею еще раз. Базовый скелет все тот же, как в хвосте у любого другого млекопитающего, но измененный, чтобы выполнять другую работу. Вернее, сам хвост - не совсем одинаков: у хвоста паукообразной обезьяны есть дополнительное количество позвонков, но сами позвонки - узнаваемо те же, что и позвонки в любом другом хвосте, включая наш собственный копчик. Вы можете вообразить себе, на что это похоже: быть обезьяной с пятью цепкими "руками" - по одной в конце каждой ноги, так же как в конце каждой руки, и хвост - на любой из которых Вы могли бы счастливо висеть? Я не могу. Но я знаю, что хвост паукообразной обезьяны гомологичен с моим копчиком, так же как чрезвычайно длинная и сильная кость крыла птеродактиля гомологична с моим мизинцем.
Вот еще один удивительный факт. Копыто лошади соответствует ногтю вашего среднего пальца руки (или ногтю вашего среднего пальца ноги). Лошади буквально ходят на цыпочках, в отличие от нас, когда мы ходим на том, что называем цыпочками. Они почти полностью потеряли другие пальцы. У лошади гомологи нашего указательного и безымянного пальца и их эквиваленты на задних ногах сохранились как крошечные грифельные косточки, соединенные с пястной/плюсневой костью среднего пальца, и невидимы снаружи шкуры. Пястная (плюсневая) кость лошади гомологична нашей средней пястной костью нашей руки (или плюсневой нашей ноги). Весь вес лошади - очень существенный в случае шайрской или клейдесдальской породы - держится на средних пальцах рук и ног. Гомологии, например, с нашими средними пальцами или таковыми у летучей мыши, полностью очевидны. Ни у кого нет сомнений относительно них; и, как будто чтобы вдолбить эту мысль в голову, иногда рождаются аномальные лошади с тремя пальцами на каждой ноге, средний из которых служит в качестве нормальной "ноги", на двух боковых есть миниатюрные копыта (см. рисунок на обороте).
Вы видите, насколько она красива, эта идея почти неограниченной модификации в течение необъятного времени, когда каждая измененная форма сохраняет несомненные следы оригинала? Я упиваюсь литоптернами, вымершими южноамериканскими травоядными животными, не связанными близким родством ни с одним из современных животных и очень отличавшимися от лошадей - за исключением того, что у них были почти идентичные ноги и копыта. У лошадей (в Северной Америке) и литоптерн (в Южной Америке, которая была в те дни гигантским островом, Панамский перешеек был делом будущего) в ходе эволюции независимо произошла совершенно одинаковая редукция всех пальцев передних и задних ног, кроме средних, и образовались идентичные копыта на кончиках пальцев. По-видимому, у травоядного млекопитающего есть не так уж и много путей стать быстрым бегуном. Лошади и литоптерны обнаруживают один и тот же путь - редуцирования всех пальцев, кроме средних - и они достигли одного и того же результата. Коровы и антилопы нашли другое решение: редуцирование всех пальцев, кроме двух.
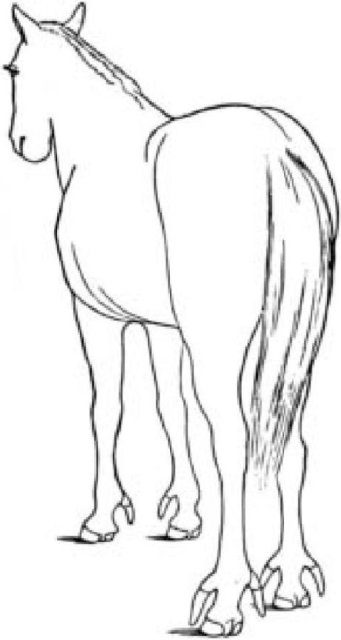
Многодактилическая лошадь
Следующее утверждение кажется парадоксальным, но можно понять, что оно логично, а также насколько оно важно как наблюдение. Скелеты всех млекопитающих идентичны, но их отдельные кости различны. Решение парадокса лежит в моем преднамеренном использовании "скелета" для совокупности костей, упорядоченно соединенных одна с другой. Форма отдельных костей, с этой точки зрения, не является свойством "скелета" вообще. "Скелет", в этом особом смысле, пренебрегает формой отдельных костей и затрагивает только порядок, в котором они соединяются: ""кость с костью своею" по словам Иезекииля и, более живо, в песне, основанной на этой фразе:
Кость вашего пальца соединена с Костью вашей ступни, Кость вашей ступни соединена с Костью вашей лодыжки, Кость вашей лодыжки соединена с Костью вашей голени, Кость вашей голени соединена с Костью вашего колена, Кость вашего колена соединена с Костью вашего бедра, Кость вашего бедра соединена с Вашей тазовой костью, Ваша тазовая кость соединена с Вашей костью спины, Ваша кость спины соединена с Вашей лопаткой, Ваша лопатка соединена с Костью вашей шеи, Кость вашей шеи соединена с Костью вашей головы, я слышу слово Бога!
Дело в том, что эту песню можно применить буквально к любому млекопитающему, безусловно, к любому наземному позвоночному животному, и гораздо более детально, чем предполагают эти слова. Например, Ваша "кость головы" или череп, содержит двадцать восемь костей, главным образом объединенных твердыми "швами", но с одной большой, подвижной костью (нижней челюстью). И замечательная вещь, что, плюс-минус случайная кость тут и там, один и тот же набор из двадцати восьми костей, которые могут быть четко обозначены одними и теми же названиями, обнаружены у всех млекопитающих.
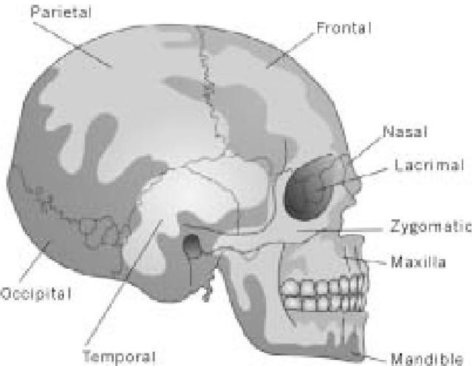
Человеческий череп
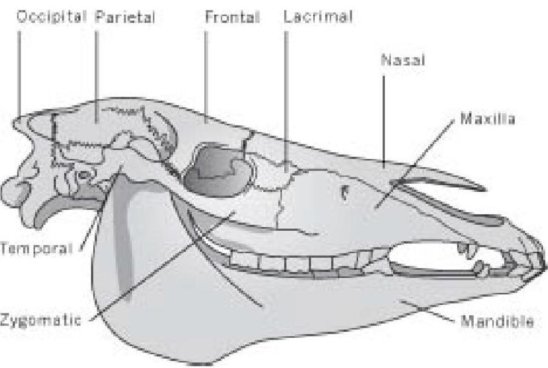
Череп лошади
Ваша кость шеи соединена с Вашей затылочной костью, Ваша затылочная кость соединена с Вашей теменной костью, Ваше темя соединено с Вашей лобной костью, Ваша лобная кость соединена с Вашей носовой костью... Ваша 27-ая кость соединена с Вашей 28-ой костью...
Все они одни и те же, независимо от факта, что форма отдельных костей радикально отличается от одного млекопитающего к другому.
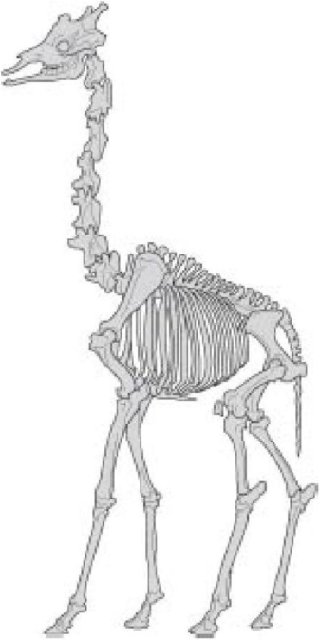
Жираф
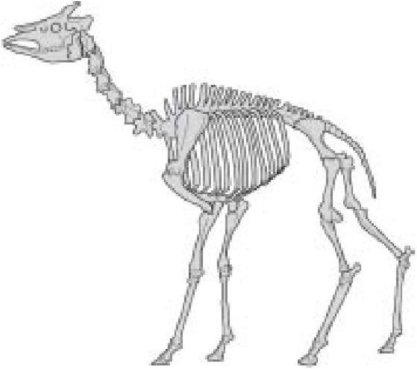
Окапи
Какой вывод мы сделаем из всего этого? Мы здесь ограничились современными животными, таким образом мы не видим эволюцию в действии. Мы - детективы, запоздало прибывшие на место события. И особенности схожести среди скелетов современных животных именно таковы, какие мы должны ожидать, если все они произошли от общего предка, одни раньше, другие позже. Предковый скелет постепенно изменялся с течением веков. Некоторые пары животных, например жирафы и окапи, объединяет недавний предок. Не очень правильно описывать жирафа как растянутого вертикально окапи, поскольку оба - современные животные. Но было бы обоснованным предположить (что, между прочим, подтверждается ископаемыми свидетельствами, но мы не говорим об ископаемых в этой главе), что их общий предок, вероятно, был больше похож на окапи, чем на жирафа. Точно так же импалы и гну - близкие кузены друг другу, и немного более дальние родственники жирафов и окапи. Все четверо -более дальние родственники другим парнокопытным, таким как свиньи и бородавочники (которые являются кузенами друг друга и пекари). Все парнокопытные - более дальние родственники лошадей и зебр (которые не имеют раздвоенных копыт и являются близкими кузенами друг друга). Мы можем продолжать сколько угодно, заключая в скобки пары кузенов в группах, и группируя группы кузенов, и (группы (групп (групп кузенов))). Я автоматически плавно перешел к использованию скобок, и Вы точно знаете, что они означают. Значение скобок в нижеследующем понятно сразу, потому что Вы уже знаете все о двоюродных братьях с общей бабушкой и дедушкой и троюродных братьях с общими прабабушкой и прадедушкой, и так далее:
{(лиса волк) (леопард лев)} {(жираф окапи) (гну импалы)}.
Все указывает на простое ветвящееся дерево происхождения -генеалогическое дерево.
Я подразумевал, что дерево подобий - на самом деле генеалогическое дерево, но напрашивается ли это заключение? Есть ли какие-нибудь альтернативные интерпретации? Ну, едва ли! Иерархическая схема подобий была замечена креационистами в додарвинистские времена, и у них действительно было неэволюционное объяснение, смущающе натянутое. Схемы подобия, на их взгляд, отражают темы в мыслях проектировщика. У него были различные идеи, как сделать животных. Его мысли вертелись вокруг темы млекопитающего, и, независимо, они вертелись вокруг темы насекомого. В теме млекопитающего идеи проектировщика были аккуратно и иерархически разделены пополам в подтемы (скажем, тема парнокопытного), и под-под-темы (скажем, тема свиньи). В этом есть весомый элемент специальных оговорок и принятия желаемого за действительное, и в настоящее время креационисты редко прибегают к этому. Действительно, как и в случае со свидетельством от географического распределения, которое мы обсуждали в последней главе, они редко обсуждают сравнительные свидетельства вообще, предпочитая придерживаться ископаемых, о которых их (неправильно) учили думать, как о многообещающей почве.
БЕЗ ЗАИМСТВОВАНИЯ
Чтобы подчеркнуть, насколько странная идея создателя, твердо придерживающегося "тем", подумайте, что любой разумный человеческий проектировщик весьма счастлив заимствовать идею одного из своих изобретений, если она принесла бы пользу другому. Возможно, есть " тема" дизайна самолета, отдельная от " темы" дизайна поезда. Но деталь самолета, скажем улучшенный дизайн для персональных лампочек над сидениями, вполне могла бы быть заимствована для использования в поездах. Почему бы и нет, если это служит одной и той же цели в обоих? Когда легковые автомобили впервые были изобретены, название "безлошадный вагон" говорило нам, откуда появилась часть вдохновения. Но гужевые транспортные средства не нуждаются в руле - Вы используете узды, чтобы править лошадьми - таким образом, у руля должен был быть другой источник. Я не знаю, откуда он взялся, но подозреваю, что он был заимствован из совершенно другой техники, техники судов. Прежде чем быть замененным рулем, который был введен приблизительно в конце девятнадцатого века, первоначальное устройство руления автомобиля было рукояткой, также заимствованной у судов, но перемещенное с зада на перед транспортного средства.
Если перья - хорошая идея в "теме" птиц, настолько, что они есть у каждой отдельной птицы без исключения, летает она или нет, почему ни у одного млекопитающего их нет? Почему проектировщик не заимствовал это остроумное изобретение, перо, хотя бы для одной летучей мыши? Ответ эволюциониста ясен. Все птицы унаследовали свои перья от их общего предка, у которого были перья. Ни одно млекопитающее не происходит от этого предка. Это так просто. Дерево подобий является генеалогическим деревом. Та же история с каждой ветвью, подветвью и под- под-ветвью дерева живого.
Теперь мы подошли к интересному моменту. Есть много красивых примеров, в которых на первый взгляд выглядит, как если идеи могли быть "заимствованы" с одной части дерева и привиты на другую, как сорт яблок, привитый на стволе. Дельфин, являющийся маленьким китом, поверхностно напоминает различных больших рыб. Одну из этих рыб, дорадо (большая корифена, Coryphaena hippurus) даже иногда называют "дельфином". Дорадо и настоящие дельфины имеют одну и ту же обтекаемую форму, подходящую для их сходного образа жизни быстрых охотников вблизи поверхности моря. Но их техника плавания, хотя на первый взгляд похожа, не была заимствована от одного другим, как Вы можете скоро увидеть, если посмотрите на детали. Хотя и те и другие получают скорость главным образом благодаря хвосту, дорадо, как все рыбы, двигает своим хвостом из стороны в сторону. А настоящий дельфин выдает свою историю млекопитающего, взмахивая хвостом вверх и вниз. Волна из-стороны-в-сторону, передающаяся вдоль предкового позвоночника рыбы, была унаследована ящерицами и змеями, которые фактически, можно сказать, "плавают" по суше. Сравните это с галопирующей лошадью или гепардом. Скорость создается благодаря изгибанию позвоночника, как у рыб и змей; но у млекопитающих позвоночник изгибается вверх и вниз, а не из стороны в сторону. Интересный вопрос, как был совершен этот переход в родословной млекопитающих. Возможно, была промежуточная стадия, которая едва сгибала свой позвоночник вообще в любом направлении, подобно лягушке. С другой стороны, крокодилы способны к галопирующей (устрашающе быстрой) походке, так же как к использованию походки, подобной ящерице, более обычной среди рептилий. Предки млекопитающих вовсе не были похожи на крокодилов, но, возможно, крокодилы показывают нам, как промежуточный предок мог комбинировать эти две походки.
Как бы то ни было, предки китов и дельфинов были стопроцентными сухопутными млекопитающими, которые, конечно, галопировали через прерии, пустыни или тундры, изгибая позвоночник вверх-вниз. И когда они вернулись в море, они сохранили свое унаследованное движение спины вверх-вниз. Если змеи "плавают" на суше, дельфины "галопируют" в море! Соответственно, хвост дельфина может поверхностно быть похожим на раздвоенный хвост дорадо, но он стоит горизонтально, тогда как хвостовой плавник дорадо расположен в вертикальной плоскости. Есть много других аспектов, в которых история дельфина написана повсюду в них, и я дойду до этого в главе с соответствующим названием.
Есть другие примеры, где поверхностное подобие настолько сильно, что кажется довольно трудно отвергнуть гипотезу "заимствования", но более близкое рассмотрение показывает, что мы должны это сделать. Животные могут выглядеть настолько подобными, что Вы ощущаете, что они должны быть связаны родством. Но затем оказывается, что сходства, хотя и впечатляющие, уступают различиям, если смотреть на все тело. "Свертывающиеся мокрицы" (см. выше) - знакомые маленькие существа с большим количеством ног, которые обычно сворачиваются в защитный шар, как броненосец. Действительно, это может быть источником латинского названия Armadillidium. Это название одной разновидности "свертывающихся мокриц", которая является ракообразным, точнее мокрицей, которая родственна креветкам, но живет на суше, где она выдает своих недавних водных предков тем, что дышит жабрами, которые нужно держать влажными. Но суть истории в том, что существует совершенно другая разновидность "свертывающихся мокриц", которая является вовсе не ракообразным, а многоножкой. Если посмотреть на них свернутыми, можно подумать, что они почти идентичны. Все же одна - измененная мокрица, в то время как другая - измененная (в том же направлении) многоножка. Если их развернуть и осмотреть тщательно, то сразу видно по крайней мере одно важное различие. У многоножки-броненосца две пары ног на большинстве сегментов, у мокрицы только один. Разве это не красиво, вся эта бесконечная модификация? Более детальный осмотр покажет, что в сотнях аспектов многоножка-броненосец действительно напоминает обычную многоножку. Сходство с мокрицей является поверхностным - конвергентным [сходящимся].
Многоножка
Мокрица
Почти любой зоолог, не являющийся специалистом, скажет, что череп на странице напротив принадлежит собаке. Специалист обнаружил бы, что это не настоящий череп собаки, отметив два заметных отверстия в небе. Они являются красноречивым признаком сумчатых, большой группы млекопитающих, в настоящее время существующих главным образом в Австралии. В действительности это череп тилацина, "тасманийского волка". Тилацины и настоящие собаки (например, динго, с которыми они конкурировали в Австралии и Тасмании) конвергентно пришли к очень схожему черепу, потому что у них (был, увы, в случае несчастного тилацина), схожий образ жизни.
Я уже упоминал великолепную фауну сумчатых млекопитающих Австралии в главе о географическом распространении животных. Вопрос, которого касается эта глава - многочисленные конвергенции между этими сумчатыми и большим спектром их аналогов среди "плацентарных" (то есть не сумчатых) млекопитающих, которые доминируют в остальном мире. Хотя далеко не идентично, даже во внешних особенностях, каждое сумчатое на рисунке на обороте достаточно схоже со своим плацентарным аналогом - то есть с плацентарным, которое близко практикует ту же самую "профессию" - достаточно схоже, чтобы поразить Вас, но, конечно, не достаточно, чтобы предполагать "заимствование" создателем.
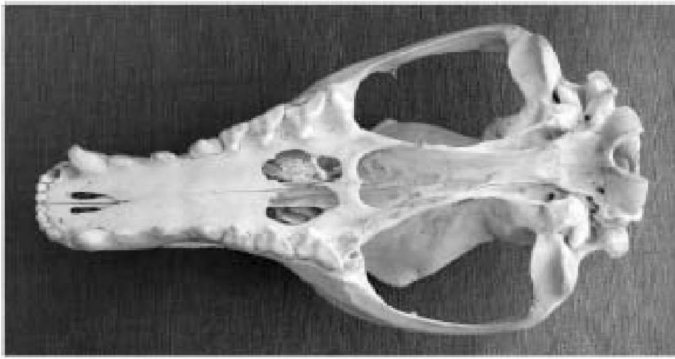
Череп «сумчатого волка» или «тасманийского тигра»
Половая перетасовка генов в генофонде может быть расценена как своего рода заимствование или совместное использование генетических "идей", но половая рекомбинация ограничена в пределах одного вида и поэтому не имеет отношения к этой главе о сравнениях между видами: например, о сравнении сумчатых и плацентарных млекопитающих. Интересно, что высокий уровень заимствования ДНК распространен среди бактерий. В процессе, иногда расцениваемом в качестве своего рода предшественника полового размножения, бактерии - даже связанные весьма отдаленным родством штаммы бактерий - обмениваются "идеями" ДНК с распутной развязностью. "Заимствование идей" -действительно один из главных способов, с помощью которых бактерии подхватывают полезные "трюки", такие как сопротивление отдельным антибиотикам.
Этот феномен часто именуют довольно бесполезным названием "трансформация". Все потому, что, когда он был обнаружен в 1928 году Фредериком Гриффитом, никто не понимал ДНК. Гриффит пришел к заключению, что невирулентный штамм стрептококка мог подхватить вирулентность от совершенно другого штамма, даже при том, что этот вирулентный штамм был мертв. В настоящее время мы сказали бы, что невирулентный штамм включил в свой геном часть ДНК от мертвого вирулентного штамма (ДНК не страшно быть "мертвой", это всего лишь закодированная информация). На языке этой главы невирулентный штамм "заимствовал" генетическую "идею" у патогенного штамма. Конечно, бактерии, заимствующие гены у других бактерий, это совсем другое дело, чем проектировщик, заимствующий свои собственные идеи из одной "темы" и вновь использующий их в другой теме. Однако, это интересно, потому что, если это было бы настолько распространено у животных, как у бактерий, это сделало бы более трудным опровержение гипотезы "заимствующего проектировщика". Что если бы летучие мыши и птицы вели себя как бактерии в этом отношении? Что если куски генома птицы могли бы быть переправлены, возможно, бактериальной или вирусной инфекцией, и внедрены в геном летучей мыши? Может быть, один вид летучей мыши мог бы внезапно вырастить перья, кодирующая перо информация ДНК была заимствована в генетическом варианте компьютерного "Копировать и Вставить".
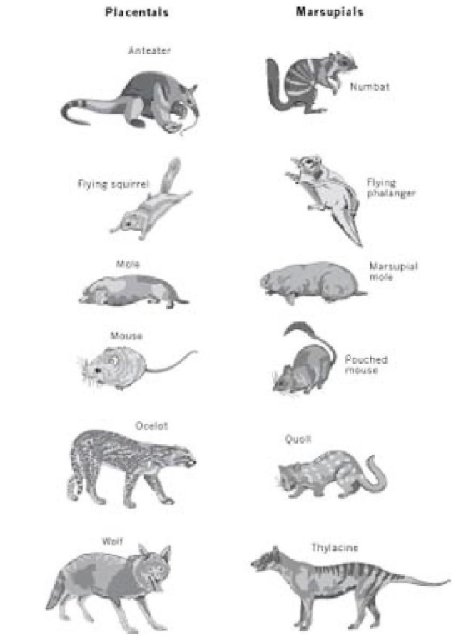
Плацентарные и сумчатые коллеги
У животных, в отличие от бактерий, передача генов, похоже, почти полностью ограничивается половыми актами в пределах вида. Действительно, вид может быть вполне удовлетворительно определен как ряд животных, которые участвуют в передаче генов между собой. Если две популяции одного вида были разделены довольно долгое время, так что они больше не могут обмениваться генами половым способом (обычно после начального периода вынужденного географического разделения, как мы видели в Главе 9), мы теперь определяем их как отдельные виды, и они никогда не будут снова обмениваться генами, исключая вмешательство человеческих генных инженеров. Мой коллега Джонатан Ходжкин, профессор генетики в Оксфорде, знает только три условных исключения к правилу, что передача генов ограничена видами: у червей нематод, у плодовых мушек, и (гораздо значительнее) у бделлоидных коловраток.
Эта последняя группа особенно интересна, потому что у них единственных среди крупных групп эукариот размножение бесполое. Могло ли случиться так, что они могли обойтись без полового размножения, потому что вернулись к древнему бактериальному способу обмениваться генами? Межвидовая передача генов, похоже, более обычна у растений. Паразитарное растение повилика (Cuscuta) служит донором генов для растения-хозяина, вокруг которого оно обвивается.
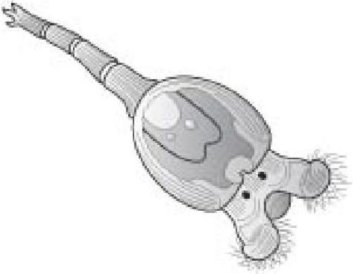
Бделлоидная коловратка
Я не определился в отношении политики ГМ-продуктов, разрываясь между потенциальными выгодами для сельского хозяйства с одной стороны и инстинктами предосторожности с другой. Но один аргумент, который я не слышал прежде, стоит кратко упомянуть. Сегодня мы ругаем то, как наши предшественники ввозили виды животных в чужие земли только ради удовольствия. Американская серая белка была завезена в Британию бывшим герцогом Бедфорда: фривольная прихоть, которую мы теперь осознаем как пагубно безответственную. Интересно задаться вопросом, не будут ли таксономисты будущего сожалеть о том, как наше поколение балуется с геномами: переносит, например, гены "антифриза" из арктической рыбы в помидоры, чтобы защитить их от мороза. Ген, дающий медузам флуоресцентный свет, был заимствован у них учеными и вставлен в геном картофеля, в надежде на то, чтобы заставить его светиться, когда он нуждается в поливе. Я даже читал о "художнике", который планирует "композицию", состоящую из люминесцентных собак, светящихся с помощью генов медузы. Такое развращение науки во имя претенциозного "искусства" оскорбляет все мои чувства. Но не идет ли вред дальше? Могут ли эти фривольные капризы подорвать обоснованность будущих исследований эволюционного родства? В действительности, я сомневаюсь в этом, но, возможно, вопрос стоит, по крайней мере, поставить на обсуждение, в духе предосторожности. Вся суть принципа предосторожности, в конце концов, в том, чтобы сейчас избежать будущих последствий выбора и действий, которые могут не быть явно опасными.
РАКООБРАЗНЫЕ
Я начал главу со скелета позвоночных, который является прекрасным примером неизменной схемы, связывающей изменчивые детали. Почти любая другая большая группа животных показала бы то же самое. Я возьму лишь еще один любимый пример: десятиногие ракообразные, группа, включающая омаров, креветок, крабов и раков-отшельников. План строения тела всех ракообразных один и тот же. Тогда как наш скелет позвоночного состоит из твердых костей внутри в остальном мягкого тела, у ракообразных экзоскелет состоит из твердых трубок, внутри которых животное содержит и защищает свои мягкие части. Твердые трубки соединены и вращаются почти таким же образом, как наши кости. Представьте, например, тонкие шарниры в лапах краба или омара и более крепкий шарнир клешни. Мускулы, приводящие в действие зажимы большого омара, находятся в трубках, составляющих клешню. Аналогичные мускулы, заставляющие человеческую руку что-либо сжимать, прикреплены к костям, проходящим внутри пальцев.
Как и позвоночные, но в отличие от морских ежей или медуз, ракообразные лево/право- симметричны, с рядом сегментов, идущих вдоль тела от головы к хвосту. Сегменты одинаковы в своем общем плане, но часто отличаются деталями. Каждый сегмент состоит из короткой трубки, которая соединена, либо жестко, либо шарниром, с двумя соседними сегментам. Как и у позвоночных, органы и системы органов ракообразного показывают повторяющуюся картину, если двигаться спереди назад. Например, главный нервный ствол, проходящий вдоль тела на брюшной стороне (не на спинной стороне, как спинной мозг позвоночного), имеет пару ганглиев (что-то вроде мини-мозга) в каждом сегменте, от которых отрастают нервы, обслуживающие сегмент. Большинство сегментов имеют по конечности с каждой стороны, каждая конечность, снова же, состоит из ряда шарнирно соединенных трубок. Конечности ракообразных обычно заканчиваются разветвлением надвое, которую во многих случаях можно назвать клешней. Голова также сегментирована, хотя, как и голова позвоночного, сегментированность здесь более замаскирована, чем в остальных частях тела. Существует пять пар конечностей, спрятанных в голове, хотя могло бы показаться немного странным назвать их конечностями, так как они изменились, став антеннами или деталями челюстного аппарата. Их поэтому обычно называют придатками, а не конечностями. Более или менее неизменно, пять сегментарных придатков головы, считая спереди, состоят из первых антенн (или антеннул), вторых антенн (часто называемых просто антеннами), нижней челюсти, первой верхней челюсти (или максиллы) и второй верхней челюсти. Антеннулы и антенны главным образом заняты опознаванием вещей. Нижние челюсти и максиллы заняты пережевыванием, размалыванием или иной обработкой пищи. Если продвигаться назад вдоль тела, сегментарные придатки или конечности довольно изменчивы, средние часто состоят из ходильных ног, в то время как те, что отрастают от последних сегментов, часто вынуждены служить другим вещам, таким как плавание.
У омара или креветки после обычных пяти сегментных придатков головы первые сегментные придатки - клешни. Следующие четыре пары - ходильные ноги. Сегменты, имеющие клешни и ходильные ноги, связаны вместе в грудь. Оставшуюся часть тела называют брюшком. Его сегменты, по крайней мере пока Вы не достигнете конца хвоста, это "плавательные ножки", перистые придатки, помогающие при плавании, весьма важные для некоторых утонченно изящных креветок. У крабов голова и грудь слились в одно большое целое, к которому прикреплены все первые десять пар конечностей. Брюшко загнуто в обратную сторону под головогрудью так, что его нельзя увидеть сверху вообще. Но если Вы перевернете краба, Вы можете ясно увидеть сегментный характер брюшка. Рисунок ниже показывает типичное узкое брюшко самца краба. Брюшко самки более широкое и напоминает передник, как его действительно называют. Рак-отшельник необычен тем, что его брюшко асимметрично (чтобы вписываться в пустую раковину моллюска, которая служит ему домом), мягкое и не покрыто панцирем (потому что раковина моллюска обеспечивает защиту).
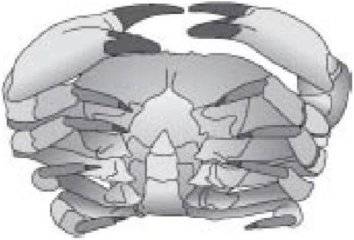
Краб мужского пола показывает узкий, назад свернутый живот
Чтобы составить представление о том, как замечательно тела ракообразных модифицированы в деталях, в то время как сам план строения тела вообще не изменен, посмотрите напротив на ряд рисунков известного зоолога девятнадцатого века Эрнста Геккеля, вероятно, самого преданного ученика Дарвина в Германии (преданность не была взаимна, но даже Дарвин, конечно, восхищался изобразительным мастерством Геккеля). Так же, как со скелетом позвоночного, посмотрите на каждую часть тела этих крабов и речного рака, и увидите, что Вы неизменно обнаружите их точные аналоги во всех остальных. Каждая часть экзоскелета соединяется с "такими же" частями, но формы самих частей очень различны. Еще раз, "скелет" является неизменным, в то время как его части - вовсе нет. И еще раз, очевидное - я бы сказал единственное разумное -объяснение в том, что все эти ракообразные унаследовали план их скелета от общего предка. Отдельных их детали отлиты в богатое разнообразие форм. Но сам план остается точно таким же, как был унаследован от предка.
ЧТО Д’АРСИ ТОМПСОН СДЕЛАЛ БЫ С КОМПЬЮТЕРОМ?
В 1917 году великий шотландский зоолог Д’Арси Томпсон написал книгу под названием "Рост и Форма", в последней главе которой он представил свой знаменитый "метод трансформации". Он чертил животное на миллиметровке, а затем искажал миллиметровку особым математически точно определимым способом и показывал, что форма первоначального животного превратилась в другое, родственное животное. Вы можете представить первоначальную миллиметровку как кусок резины, на котором Вы рисуете свое первое животное. Потом преобразованная миллиметровка была бы эквивалентна тому же куску резины, растянутому или деформированному некоторым определенным математическим способом. Например, он взял шесть видов крабов и нарисовал одного из них, Geryon, на обычной миллиметровке (недеформированный лист резины). Затем он исказил свой математический "резиновый лист" пятью отдельными способами, чтобы достичь приблизительного изображения других пяти видов крабов. Математические детали не имеют значения, хотя они и восхитительны. Вы можете ясно видеть, что нужно не много, чтобы трансформировать одного краба в другой. Сам Д’Арси Томпсон не очень интересовался эволюцией, но нам легко представить, что должны были бы сделать генетические мутации, чтобы вызвать изменения подобные этим. Это не означает, что мы должны считать Geryon или любого другого из этих шести крабов предком других. Ни один из них им не был, и, в любом случае, дело не в этом. Дело в том, что независимо от того, на кого был похож предковый краб, преобразования подобного рода могли изменить любой из этих шести видов (или предполагаемого предка) в любой другой.

Ракообразные Хекеля. Эрнст Хекель был выдающимся немецким зоологом и превосходным художником-зоологистом.
Эволюция никогда не происходит через трансформацию одной взрослой формы в другую. Вспомните, что каждый взрослый растет как эмбрион. Отбираемые мутации работали в развивающемся эмбрионе, изменяя скорость роста частей тела относительно других частей. В Главе 7 мы представили эволюцию человеческого черепа как ряд изменений в скорости роста некоторых частей относительно других частей, которыми управляют гены в развивающемся эмбрионе. Следует ожидать поэтому, что, если мы нарисуем человеческий череп на листе "математической резины", должна быть возможность деформировать эту резину некоторым математически изящным способом и достичь приблизительного сходства с черепом близкого кузена, такого как шимпанзе, или - вероятно, с большими искажениями - более дальнего родственника, такого как бабуин. И это именно то, что показал Д’Арси Томпсон. Снова же, заметьте, что это было произвольное решение нарисовать первым человеческий череп, а затем преобразовать его в череп шимпанзе и бабуина. Он мог бы с тем же успехом нарисовать, скажем, шимпанзе первым, а затем проработать необходимые искажения, чтобы создать череп человека и бабуина. Или, что более интересно для книги по эволюции, каковой не являлась его книга, он мог нарисовать, скажем, череп австралопитека сначала на недеформированной резине и вычислить, как ее преобразовать, чтобы сделать череп современного человека. Это, конечно, сработало бы точно так же, как и рисунки выше, но и имело бы эволюционное значение в более прямом смысле.
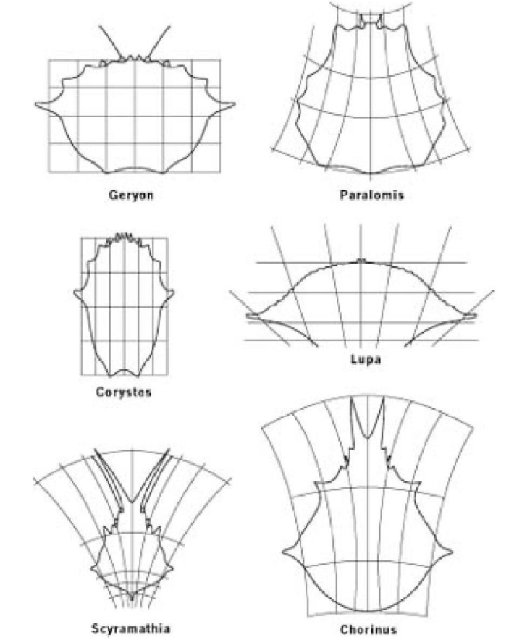
Д'Арси Томпсоновские «преобразования» краба
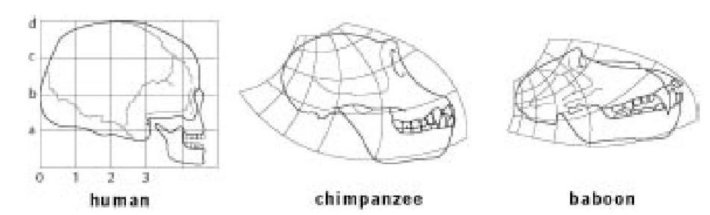
Д'Арси Томпсоновское «преобразование» черепа
В начале этой главы я представил идею "гомологии", используя руки летучих мышей и людей в качестве примера. Потворствуя идиосинкретическому использованию языка, я сказал, что скелеты были идентичны, в то время как кости были различны. Преобразования Д’Арси Томпсона предоставляют нам способ сделать эту идею более четкой. Согласно этой формулировке, два органа - например, рука летучей мыши и рука человека - являются гомологичными, если можно нарисовать один на листе резины, а затем, деформировав резину, получить другой. У математиков есть для этого слово: "гомеоморфный".
В додарвиновские времена зоологи признавали гомологию, и предэволюционисты описывали, скажем, крылья летучей мыши и человеческие руки как гомологичные. Если бы они достаточно знали математику, то были бы счастливы использовать слово "гомеоморфные". В постдарвиновские времена, когда стало общепризнано, что летучие мыши и люди разделяют общего предка, зоологи начали определять гомологию в эволюционных терминах. Гомологичные подобия - подобия, унаследованные от общего предка. Слово "аналогичный" стало использоваться для подобий, обязанных общей функции, а не родословной. Например, крыло летучей мыши и крыло насекомого были бы описаны как аналогичные, в отличие от гомологичных крыла летучей мыши и человеческой руки. Если мы хотим использовать гомологию как свидетельство факта эволюции, мы не можем использовать эволюцию для ее определения. Поэтому для этой цели удобно вернуться к предэволюционному определению гомологии. Крыло летучей мыши и человеческая рука гомеоморфны: Вы можете преобразовать одно в другое, деформируя резину, на которой оно нарисовано. Вы не можете преобразовать крыло летучей мыши в крыло насекомого таким же способом, потому что нет соответствующих частей. Широко распространенное существование гомеоморфизмов, которые определенны не в терминах эволюции, может использоваться в качестве свидетельства эволюции. Нетрудно понять, как могла эволюция поработать над рукой любого позвоночного, чтобы преобразовать ее в руку любого другого позвоночного, просто изменяя относительные скорости роста у эмбриона.
С той поры, как я начал знакомиться с компьютерами, будучи аспирантом в 1960-ых, я задался вопросом, что Д’Арси Томпсон мог бы сделать с помощью компьютера. Вопрос стал насущным в 1980ых, когда доступные компьютеры с экранами (в отличие от просто бумажных принтеров) стали распространенными. Рисование на растягиваемой резине, а затем искажение поверхность рисунка математическим способом - это просто умоляло о компьютерной обработке! Я рекомендовал Оксфордскому университету предложить грант, чтобы нанять программиста с целью запустить преобразования Д’Арси Томпсона на экране компьютера и сделать их доступными в смысле легкости в использовании. Мы получили деньги и наняли Уилла Аткинсона, первоклассного программиста и биолога, который стал моим другом и советчиком в моих собственных проектах программирования. Как только он решил трудную проблему программирования богатого набора математических искажений "резины", для него было относительно простой задачей включить это математическое колдовство в программу искусственного отбора в стиле биоморфов, подобно моим собственным программам "биоморфов", описанным здесь в Главе 2. Как и в моих программах, "игрока" сажали перед экраном, полным животных форм, и предлагали выбрать одного из них для " разведения" , поколение за поколением. Снова были "гены", которые сохранялись в течение поколений, и снова гены влияли на форму "животных". Но в данном случае способом, которым гены влияли на форму животных, было управление искажением "резины", на которой была нарисована форма животного. И потому теоретически должно было быть возможным начать, скажем, с черепа австралопитека, нарисованного на недеформированной "резине", и вывести Ваш путь среди существ со все более увеличивающимися черепами и все более короткими мордами - все более подобных человеку, другими словами. На практике оказалось очень сложно сделать нечто подобное, и я думаю, что этот факт сам по себе интересен.
Я предполагаю, одна причина, по которой это было сложно, в очередной раз, была в том, что преобразования Томпсона Д’Арси изменяют одну взрослую форму в другую взрослую форму. Как я подчеркнул в Главе 8, это не то, как работают гены в эволюции. У каждого отдельного животного есть история развития. Оно начинает как эмбрион и вырастает, за счет непропорционального роста различных частей тела, во взрослое. Эволюция - не генетически управляемое искажение одной взрослой формы в другую; это -генетически управляемое изменение в программе развития. Джулиан Хаксли (внук Томаса Генри и брат Олдоса) распознал это, когда вскоре после публикации первого издания книги Томпсона Д’Арси модифицировал "метод преобразований", чтобы изучить путь, которым ранние эмбрионы превращаются в более поздние или во взрослых. Это все, что я хочу сказать здесь о методе преобразований Д’Арси Томпсона. Я вернусь к этой теме в заключительной главе, чтобы выразить смежный момент.
Сравнительные свидетельства, как я предположил в начале этой главы, говорят даже более настойчиво, чем ископаемые свидетельства, в пользу факта эволюции. Сам Дарвин придерживался подобной точки зрения в конце своей главы в "Происхождении видов" "Взаимное родство органических существ":
Наконец, различные группы фактов, рассмотренные в этой главе, по-моему, столь ясно указывают, что бесчисленные виды, роды и семейства, населяющие земной шар, произошли каждый в пределах своего класса или группы от общих предков и затем модифицированы в процессе наследования, что я без колебаний принял бы этот взгляд, если бы даже он не был подкреплен другими фактами или аргументами.
МОЛЕКУЛЯРНЫЕ СРАВНЕНИЯ
Чего не знал, не мог знать, Дарвин, что сравнительные свидетельства становится даже более убедительным, когда мы включаем молекулярную генетику, в дополнение к анатомическим сравнениям, имевшимся в его распоряжении.
Так же, как скелет позвоночного неизменен для всех позвоночных, в то время как отдельные кости отличаются, и так же, как экзоскелет ракообразного неизменен для всех ракообразных, в то время как отдельные "трубки" варьируются, так же и код ДНК неизменен для всех живых существ, в то время как сами отдельные гены варьируются. Это - действительно поразительный факт, который показывает яснее чем что-либо еще, что все живые существа происходят от одного предка. Не только сам генетический код, но и вся генно/протеиновая система для поддержания жизни, с которой мы имели дело в Главе 8, одна и та же во всех животных, растениях, грибах, бактериях, археях и вирусах. Изменяется то, что написано кодом, не сам код. И когда мы сравниваем то, что написано кодом -реальные генетические последовательности всех этих различных существ - мы обнаруживаем своего рода иерархическое дерево подобий. Мы обнаруживаем одно и то же генеалогическое дерево -хотя намного более тщательно и убедительно детализованное - как мы делали со скелетом позвоночного, скелетом ракообразного, и, на самом деле, всей структурой анатомических подобий во всех царствах живого мира.
Если мы хотим высчитать, насколько близко связана любая пара видов - скажем, насколько близко еж к обезьяне - идеально было бы рассмотреть полные молекулярные тексты каждого гена обоих видов и сравнить до мельчайших подробностей, как ученый-библеист мог бы сравнить два свитка или фрагмента "Книги Пророка Исайи". Но это отнимает много времени и дорого. Проект "Геном человека" занял приблизительно десять лет, что соответствует многим человеко-столетиям. Хотя сейчас можно было бы достигнуть того же результата за долю того времени, это все еще будет большое и дорогостоящее предприятие, каким был бы и проект "Геном ежа". Как посадка на Луну "Аполлона" и как Большой Адронный Коллайдер (который был только что запущен в Женеве, как раз когда я пишу - гигантский размах этих международных усилий растрогал меня до слез при посещении), полная расшифровка человеческого генома - одно из тех достижений, которое заставляет меня гордиться, что я человек. Я рад, что теперь успешно завершен проект "Геном шимпанзе" , и аналогично для различных других видов. Если существующий темп прогресса продолжится (см. "Закон Ходжкика" ниже), то скоро станет экономически осуществимо секвенировать геном каждой пары видов, чью близость родства мы бы захотели измерить. Пока, однако, по большей части мы должны прибегать к выборочному обследованию определенных частей их геномов, и оно работает вполне прилично.
Мы можем брать образцы нескольких выбранных генов (или белков, последовательности которых непосредственно транслированы с генов) и сравнивать их среди разных видов. Я подойду к этому через мгновение. Но есть другие способы взять своего рода грубую, автоматическую пробу, и технологии, чтобы это делать доступным уже более продолжительное время. Ранний метод, работающий удивительно хорошо, использует иммунную систему кроликов (фактически можно использовать любое животное, какое Вам нравится, но кролики замечательно справляются с этой работой). Как часть естественной защиты тела против болезнетворных микроорганизмов, иммунная система кролика производит антитела против любого инородного белка, поступающего в кровоток. Так же, как Вы могли бы сказать, что у меня был коклюш, глядя на антитела в моей крови, так же Вы можете сказать, чему подвергался кролик в прошлом, глядя на его иммунный ответ в настоящее время. Нынешние антитела в кролике составляют историю естественных потрясений, которую наследует его плоть - включая искусственно вводившиеся белки. Если Вы введете, скажем, белок шимпанзе в кролика, то антитела, которые кролик произведет, впоследствии атакуют тот же самый белок, если он будет введен снова. Но предположим, что Ваша вторая инъекция содержит эквивалентный белок, не шимпанзе, а гориллы? Кролик, предварительно подвергнутый белку шимпанзе, будет частично вооружен против версии гориллы, но реакция будет более слабой. И он также будет вооружен против версии белка кенгуру, но реакция будет еще более слабой, учитывая, что кенгуру связан намного менее близким родством с подготовившим его шимпанзе, чем горилла. Сила иммунного ответа кролика на последующую инъекцию белка является мерой подобия этого белка с оригиналом, которому был первоначально подвергнут кролик.
Именно этим методом, используя кроликов, Винсент Сарич и Аллан Уилсон из Калифорнийского Университета в Беркли продемонстрировали в 1960-ых, что люди и шимпанзе связаны намного более близким родством друг с другом, чем кто-либо ранее себе представлял. Есть также методы, которые используют сами гены, непосредственно сравнивая их среди видов, вместо того чтобы сравнить белки, которые они кодируют. Один из самых старых и самый эффективный из этих методов называют гибридизацией ДНК. Гибридизация ДНК обычно стоит за теми высказываниями, которые можно часто услышать, типа: "У людей и шимпанзе общие 98 процентов их генов". Между тем, есть некоторая путаница, как раз относительно того, что имеется в виду под числами процентов, такими как эти. Девяносто восемь процентов чего идентично? Точное число зависит от того, насколько большими единицами мы считаем. Простая аналогия ясно дает это понять, и она делает это интересным способом, поскольку различия между аналогией и реальной вещью являются столь же показательными, как и сходства. Предположим, что у нас есть два варианта одной и той же книги, и мы хотим их сравнить. Скажем, это "Книга пророка Даниила", и мы хотим сравнить каноническую версию с древним свитком, который был только что обнаружен в пещере на берегу Мертвого моря. Сколько процентов глав двух книг идентично? Вероятно, ноль, для этого нам требуется только одно несоответствие где-нибудь во всей главе, чтобы сказать, что эти две не идентичны. Какой процент их предложений идентичен? Процент теперь будет намного выше. Еще выше будет процент идентичных слов, потому что у слов меньше букв, чем у предложений - меньше возможностей разрушить идентичность. Но подобие слова все еще нарушается, если любая из букв в слове отличается. Поэтому, если Вы выровняете рядом два текста и сравните их буква за буквой, процент идентичных букв будет еще выше, чем процент идентичных слов. Таким образом, оценка типа "98 процентов общего" ничего не означает, если мы не определим размер единиц, которые мы сравниваем. Считаем ли мы главы, слова, буквы или что? И то же самое верно, когда мы сравниваем ДНК двух видов. Если сравнивать целые хромосомы, процент - ноль, потому что требуется лишь одно крошечное различие где-нибудь на протяжении хромосом, чтобы определить хромосомы как различные.
Часто упоминаемое число приблизительно 98 процентов общего генетического материала людей и шимпанзе фактически не относится ни к числу хромосом, ни к числу целых генов, а к числу "букв" ДНК (технически пар оснований), идентичных друг другу в соответствующих генах человека и шимпанзе. Но есть подвох. Если Вы производите выравнивание наивно, то недостающая буква (или добавленная), в отличие от ошибочной буквы, приведет к несоответствию всех последующих букв, потому что все они тогда будут сдвинуты на шаг, опережая одна другую (пока не будет ошибки в противоположном направлении, чтобы снова вернуть их к синхронности). Несомненно, несправедливо позволять оценке несоответствий быть преувеличенной таким образом. Глаз ученого, просматривая два свитка "Книги пророка Даниила", автоматически справляется с этим трудно переводимым в цифры способом. Как мы можем сделать это с ДНК? Здесь мы оставим нашу аналогию с книгами и свитками и перейдем прямо к самой вещи, потому что оказывается саму вещь - ДНК - легче понять, чем аналогию!
Если Вы постепенно нагреваете ДНК, наступает момент - где-то около 85°C - когда связь между двумя нитями двойных спиралей разрывается, и две спирали разделяются. Вы можете представить 85°C, или неважно какую получившуюся температуру, как "точку плавления". Если Вы позволите ей снова остыть, каждая одинарная спираль спонтанно снова соединится с другой одинарной спиралью или фрагментом одинарной спирали, везде, где найдет фрагмент, с которым она может соединиться, используя обычные для двойной спирали правила комплементарности пар оснований. Можно подумать, что это всегда будет партнер, от которого она только что отделилась и которому, конечно, она полностью соответствует. Действительно так может быть, но обычно все не столь гладко. Фрагменты ДНК найдут другие фрагменты, с которыми они могут соединиться, и это обычно не будут в точности их первоначальные партнеры. И действительно, если Вы добавите разделенные фрагменты ДНК другого вида, фрагменты одинарных нитей вполне способны соединиться с фрагментами одинарных нитей неправильного вида, точно так же, как они соединятся с одиночными нитями правильного вида. Почему бы и нет? Это замечательное следствие переворота в молекулярной биологии Крика и Уотсона, что ДНК - всего лишь ДНК. Она не "заботится" о том, является ли она человеческой ДНК, ДНК шимпанзе или ДНК яблока. Фрагменты охотно соединяются с комплементарными фрагментами везде, где они их находят. Однако прочность связи не всегда одинакова. Однонитевые куски ДНК связываются сильнее с соответствующей одинарной нитью, чем с менее подобными одинарными нитями. Причина в том, что больше "букв" ДНК ("оснований" Уотсона и Крика) оказываются напротив партнеров, с которыми они не могут соединиться. Связь между нитями поэтому ослаблена - как у застежки-молнии с недостающими некоторыми зубьями.
Как нам измерить эту прочность связи, после того, как фрагменты от различных видов нашли друг друга и объединились? До смешного простым методом. Мы определяем "точку плавления" связей. Помните, я говорил, что точка плавления двухспиральной ДНК около 85°C. Это верно для нормальной, должным образом согласующейся двухспиральной ДНК, как, например, когда нить человеческой ДНК "отплавлена" от комплементарной нити человеческой ДНК. Но когда связь слабее - как например, когда человеческая нить соединяется с нитью шимпанзе - достаточно немного более низкой температуры, чтобы разорвать эту связь. И когда человеческая ДНК связывается с ДНК более дальнего родственника, такого как рыба или жаба, достаточно еще более низкой температуры, чтобы их разделить. Различие между точкой плавления, когда одна нить связана к другой нитью своего собственного вида, и точкой плавления, когда она связана с нитью другого вида, является нашей мерой генетического расстояния между двумя видами. Как практическое правило, уменьшение на каждый 1° Цельсия "точки плавления" приблизительно равноценно снижению на 1 процент количества соответствий букв ДНК (или увеличению на 1 процент числа недостающих зубьев в застежке-молнии).
В этом методе есть осложнения, в которые я не вдавался, и хитрые проблемы, для которых есть изобретательные решения. Например, если смешать ДНК человека с ДНК шимпанзе, то большая часть фрагментированной человеческой ДНК соединится с другими фрагментами человеческой ДНК, а большая часть ДНК шимпанзе соединится со своей же. Как отделить гибридную ДНК, чью "точку плавления" Вы действительно хотите определить, от "однородной" ДНК? Ответом является умная уловка, предусматривающая предварительную радиоактивную маркировку. Но эти детали увели бы нас слишком далеко от нашего пути. Суть здесь в том, что гибридизация ДНК - это техника, которая приводит ученых к цифрам, вроде 98 процентов генетического сходства между людьми и шимпанзе, и она приводит к очевидно более низким процентам, если перейти к парам животных, связанных более отдаленным родством.
Новейший метод измерения подобия между парой соответствующих генов различных видов является наиболее прямым и самым дорогим: фактически чтением последовательности букв непосредственно в генах, используя те же методы, которые применялись в проекте "геном человека". Хотя все еще дорого сравнивать весь геном, можно получить хорошее приближение, сравнивая выборку генов, и это теперь делается все чаще.
Какой бы ни была техника, которую мы используем для измерения подобия между двумя видами, будь то антитела кролика, или точки плавления, или прямое секвенирование, следующий шаг в значительной степени одинаков. Получив отдельное число, отображающее подобие между каждой парой видов, мы затем заносим эти числа в таблицу. Возьмите ряд видов и напишите их названия в одном и том же порядке, как во главе колонок, так и во главе строк. Затем занесите проценты подобия в соответствующие ячейки. Таблица будет треугольной (половиной квадрата), потому что, например, процент подобия между человеком и собакой будет таким же, как подобие между собакой и человеком. Так, если заполнить во всей квадратной таблице каждую из этих двух половин, любая сторона диагонали зеркально отразила бы другую.
Теперь, каких результатов нам следует ожидать? Согласно эволюционной модели можно прогнозировать, что более высокая оценка будет помещена в ячейку, соединяющую человека и шимпанзе; более низкая - в ячейку, соединяющую человека и собаку. В ячейке человека/собаки должна теоретически быть оценка, идентичная подобию в ячейке шимпанзе/собаки, потому что у людей и шимпанзе совершенно одинаковая степень родственных отношений с собаками. Она также должна быть идентичной и ячейкам обезьяны/собаки и лемура/собаки. Ведь люди, шимпанзе, обезьяны и лемуры, все связаны с собакой через их общего предка, древнего примата (который, вероятно, немного был похож на лемура). Та же самая оценка должна обнаружиться в ячейках человека/кошки, шимпанзе/кошки, обезьяны/кошки и лемура/кошки, потому что кошки и собаки связаны со всеми приматами через общего предка всех хищных. Должна быть намного более низкая оценка - в идеале одинаково низкая - во всех ячейках, объединяющих, скажем, кальмара с любым млекопитающим. И не должно иметь значения, какое выбрано млекопитающее, так как все они одинаково отдалены от кальмара.
Таковы строгие теоретические ожидания, и нет никакой причины, почему на практике они не должны были нарушаться. Если бы они были нарушены, то это было бы свидетельством против эволюции. То, что происходит фактически, оказывается - в пределах статистической погрешности - именно тем, что мы должны ожидать при условии, что эволюция имела место. Это все равно, что сказать, что, если нанести генетические расстояния между парами видов на ветви дерева, все удовлетворительно складывается. Конечно, складывается не совсем безупречно. Числовые расчеты в биологии редко реализуются с более чем приблизительной точностью.
Свидетельства от сравнения ДНК (или белка) может быть использовано, чтобы решить - исходя из эволюционного предположения - какие пары животных являются более близкими кузенами, чем другие. Что превращает их в чрезвычайно сильное свидетельство эволюции - так это возможность построить дерево генетических подобий отдельно для каждого гена поочередно. И важный результат состоит в том, что каждый ген приводит к приблизительно тому же дереву живого. Еще раз, это именно то, что ожидалось бы, если бы Вы имели дело с истинным генеалогическим деревом. Это не то, что ожидалось бы, если бы проектировщик оглядел весь животный мир и выискал и выбрал - или "заимствовал" - лучшие белки для работы, везде, где бы они ни находились в животном мире.
Самое раннее крупномасштабное исследование в этом направлении было проведено группой генетиков в Новой Зеландии во главе с профессором Дэвидом Пенни. Группа Пенни взяла пять генов, которые, хотя и не идентичны у всех млекопитающих, были достаточно подобны, чтобы заслужить у всех одно и то же название. Детали не имеют значения, но, во избежание недоразумений, эти пять генов были генами гемоглобина A, гемоглобина B (гемоглобины придают крови ее красный цвет), фибринопептида A, фибринопептида B (фибринопептиды используются в свертывании крови), и цитохрома C (который играет важную роль в клеточной биохимии). Они выбрали одиннадцать млекопитающих для сравнения: макаку-резус, овцу, лошадь, кенгуру, крысу, кролика, собаку, свинью, человека, корову и шимпанзе.
Пенни и его коллеги размышляли статистически. Они хотели вычислить вероятность того, что по чистой случайности две молекулы привели бы к одному и то же генеалогическому дереву, если бы эволюция не была реальностью. Таким образом, они попытались представить себе все возможные деревья, которые могли закончиться одиннадцатью потомками. Это удивительно большое число. Даже если Вы ограничитесь "двоичными деревьями" (то есть деревьями с ветвями, которые разделяются только надвое - а не натрое или больше), общее количество возможных деревьев составит больше чем 34 миллиона. Ученые терпеливо нашли каждое из этих 34 миллионов деревьев и сравнили каждое с каждым из остальных 33 999 999 деревьев. Нет, конечно они этого не делали! Это заняло бы слишком много компьютерного времени. Однако они разработали разумное статистическое приближение, сокращенный аналог этих титанических расчетов.
Вот как этот метод приближения работает. Они взяли первый из тех пяти генов, скажем, гемоглобин-A (во всех случаях я использую название белка, чтобы обозначить ген, который кодирует этот белок). Из всех тех миллионов деревьев они хотели найти наиболее "экономное" в отношении гемоглобина-A. Экономное здесь означает "требующее предположить минимальное число эволюционных изменений". Например, все те тысячи деревьев, предполагавших, что наиболее близкий кузен человека был кенгуру, в то время как люди и шимпанзе связаны более отдаленным родством, оказались очень неэкономными деревьями: они должны были предположить множество эволюционных изменений, чтобы привести к результату, что у кенгуру и людей был недавний общий предок. Вердикт гемоглобину-A был в таком духе:
Это ужасно неэкономное дерево. Мало того, что нужно было бы выполнить много мутационной работы, чтобы закончить со столькими различиями в людях и кенгуру, несмотря на наше близкое родство, предполагаемому этим деревом, также нужно было бы проделать много мутационной работы в другом направлении, чтобы гарантировать, что, несмотря на их большое расхождение на этом конкретном дереве, люди и шимпанзе каким-то образом оказались обладателями столь схожего гемоглобина-A. Я голосую против этого дерева.
Гемоглобин-A выносит вердикты такого рода, некоторые более благоприятные, чем другие, каждому из этих 34 миллионов деревьев, и, наконец, приходит к выбору нескольких дюжин деревьев высокого ранга. О каждом из этих высокоранговых деревьев гемоглобин-A мог бы сказать примерно так:
Это дерево определяет людей и шимпанзе как близких кузенов, и оно определяет овец и коров как близких кузенов, и оно помещает кенгуру на отдельную ветвь. Оно оказалось очень хорошим деревом, потому что почти не заставляет делать какую-либо мутационную работу вообще, чтобы объяснить эволюционные изменения. Это -на диво экономное дерево. Оно получило голос гемоглобина-A!
Конечно, было бы хорошо, если бы гемоглобин-A и каждый другой ген могли бы предложить единственное дерево максимальной экономичности, но мы слишком многого хотим. Среди этих 34 миллионов деревьев ожидается только, что несколько слегка различных деревьев должны конкурировать за высокий рейтинг по гемоглобину-A.
А как насчет гемоглобина-B? Как насчет цитохрома-C? Каждый из этих пяти белков имеет право на свое собственное отдельное голосование, чтобы выявить свои собственные предпочтительные (то есть наиболее экономные) деревья из числа тех 34 миллионов деревьев. Для цитохрома-C было бы вполне возможно прийти к совершенно другим результатам голосования за наиболее экономное дерево. Могло бы оказаться, что цитохром-C людей действительно очень похож на цитохром-C кенгуру и очень отличается от такового у шимпанзе. Далеко не приветствующий соединение в близкую пару овец и коров, обнаруженную гемоглобином-A, цитохром-C мог бы обнаружить, что едва ли нужны мутации вообще, чтобы разместить овец очень близко, скажем, к обезьянам, и чтобы разместить коров очень близко к кроликам. Согласно гипотезе творения, нет никакой причины, почему этого не должно происходить. Но что фактически обнаружили Пенни и его коллеги, это удивительно высокое согласование среди всех пяти белков (и они использовали еще более умную статистику, чтобы показать, насколько маловероятно случайное возникновение такого согласования). Все пять белков "голосовали" практически за одно и то же подмножество деревьев из числа 34 миллионов возможных деревьев. Это, конечно, именно то, что мы ожидали бы при условии, что действительно есть только одно истинное дерево, связывающее все одиннадцать животных, и что это - генеалогическое дерево: дерево эволюционного родства. Кроме того, это дерево согласия, за которое голосовали все эти пять молекул, оказалось тем же, которое зоологи уже вывели на анатомических и палеонтологических, не молекулярных, основаниях.
Исследование Пенни было опубликовано в 1982 году, довольно недавно. Прошедшие годы видели обширное увеличение подробных свидетельств на основании точных последовательностей генов большого числа видов животных и растений. Согласованность по большинству экономных деревьев теперь простирается далеко за пределы одиннадцати видов и пяти молекул, которые изучал Пенни и его коллеги. Их исследование было только хорошим примером, очень сильным, как доказали их статистические свидетельства. Общая сумма информации о генетических последовательностях, доступных теперь, не оставляет по этому вопросу никаких мыслимых сомнений. Намного более убедительные, даже чем (также очень убедительные) ископаемые свидетельства, свидетельства от сравнений генов сходятся, быстро и решительно, на единственном великом дереве живого. Выше помещено дерево для одиннадцати видов исследования Пенни, которое представляет собой современное всеобщее голосование многих различных частей генома млекопитающих. Устойчивость согласованности между всеми различными генами в геноме вселяет в нас уверенность не только в исторической точности самого консенсусного дерева, но также в факт, что эволюция имела место.
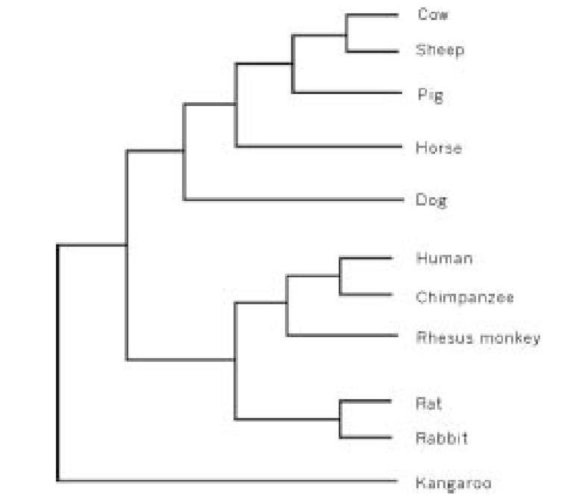
Фамильное древо для 11 разновидностей Пенни
Если молекулярная генетическая технология продолжит расширяться с ее нынешней экспоненциальной скоростью, то году к 2050 получить полную последовательность генома животного будет дешево и быстро, едва ли не труднее, чем измерить его температуру или его кровяное давление. Почему я говорю, что генетическая технология развивается по экспоненте? Может мы даже могли бы ее измерить? Есть аналог с компьютерной технологией, названный Законом Мура. Названный в честь Гордона Мура, одного из основателей компании компьютерных микросхем Intel, он может быть выражен различными способами, потому что несколько показателей компьютерной мощности связаны друг с другом. Один вариант закона утверждает, что количество модулей, которые могут быть упакованы в интегральную схему заданного размера, удваивается за срок от восемнадцати месяцев до двух лет или около того. Это - эмпирический закон, что означает, что он не выводится из некоторой теории, а просто оказывается действительным, когда Вы оцениваете данные. Он справедлив уже в течение приблизительно пятидесяти лет, и многие эксперты думают, что так будет в течение по крайней мере еще нескольких десятилетий. Другие экспоненциальные тенденции с подобным удвоением во времени, которые могут быть расценены как версии Закона Мура, включают увеличение скорости вычислений и размера памяти в расчете на единицу стоимости. Экспоненциальные тенденции всегда приводят к потрясающим результатам, как продемонстрировал Дарвин, когда при помощи своего сына, математика Джорджа, он взял слона как пример медленно размножающегося животного и показал, что лишь через несколько столетий неограниченного экспоненциального роста потомки всего одной пары слонов покрыли бы землю. Само собой разумеется, рост популяции слонов на практике не экспоненциален. Он ограничен конкуренцией за пищу и пространство, болезнями и многим другим. Это действительно было самым главным для Дарвина, поскольку здесь вступает естественный отбор.
Но Закон Мура действительно остается в силе, по крайней мере приблизительно, уже в течение пятидесяти лет. Хотя ни у кого нет особо четкого представления почему, различные показатели компьютерной мощности на практике фактически увеличивались по экспоненте, тогда как тенденция слона Дарвина экспоненциальна только теоретически. Мне пришло на ум, что может существовать подобный закон в отношении генетической технологии и секвенирования ДНК. Я подсказал это Джонатану Ходжкину, оксфордскому профессору генетики (который когда-то был моим студентом-воспитанником). К моему восхищению оказалось, что он уже думал об этом - и измерил это при подготовке к лекции в своей старой школе. Он оценил стоимость секвенирования ДНК стандартной длины для четырех дат в истории, 1965, 1975, 1995 и 2000 года. Я перевел его числа в "отдачу от вложенных средств", или в то, "сколько ДНК можно секвенировать за 1 000 £?" Я подготовил иллюстрации в логарифмическом масштабе, выбранном, потому что экспоненциальная тенденция всегда выявляется как прямая линия, когда она построена в логарифмическом масштабе. Достаточно уверенно четыре точки Ходжкина довольно хорошо ложатся на прямую линию. Я построил линию по этим точкам (по технике линейной регрессии, см. примечание на стр. 112), а затем взял на себя смелость спроецировать ее на будущее. Позже, как раз когда эта книга собиралась в печать, я показал этот раздел профессору Ходжкину, и он сообщил мне новые данные, которые он знал: геном утконоса, который был секвенирован в 2008 году (утконос - хорошая кандидатура из-за своего стратегического положения на дереве живого: его общий с нами предок, жил 180 миллионов лет назад, что почти в три раза раньше вымирания динозавров). Я нарисовал точку утконоса как звездочку на графике, и можно видеть, что она вполне точно ложится рядом с намеченной линией, которая была вычислена на основании более ранних данных.
Наклон линии для того, что я теперь называю (без разрешения) Законом Ходжкина, только немного меньше, чем для Закона Мура. Время удвоения немного больше, чем два года, тогда как время удвоения в Законе Мура немного меньше двух лет. Технология ДНК сильно зависит от компьютеров, таким образом, будет хорошим предположением, что Закон Ходжкина, по крайней мере частично, зависит от Закона Мура. Стрелки справа указывают размеры генома различных существ. Если следовать за стрелкой влево, пока она не достигнет наклонной линии Закона Ходжкина, можно прочесть оценку, когда будет возможно секвенировать геном такого же размера как у рассматриваемого существа всего за 1 000 £ (по сегодняшним деньгам). Для генома размером с геном дрожжей мы должны ожидать всего лишь приблизительно до 2020 года. Для нового генома млекопитающего (насколько можно судить по таким примерным вычислениям, все млекопитающие требуют одинаковых затрат), предполагаемая дата - чуть ранее 2040 года. Это волнующая перспектива: массивная база данных последовательностей ДНК, полученная дешево и легко из всех уголков животного и растительного царств. Подробное сравнение ДНК заполнит все пробелы в нашем знании о фактическом эволюционном родстве каждого вида с любым другим: мы узнаем с полной уверенностью все генеалогическое дерево всех живых существ. Кто знает, как мы его начертим; оно не впишется ни в какой лист бумаги практичного размера.
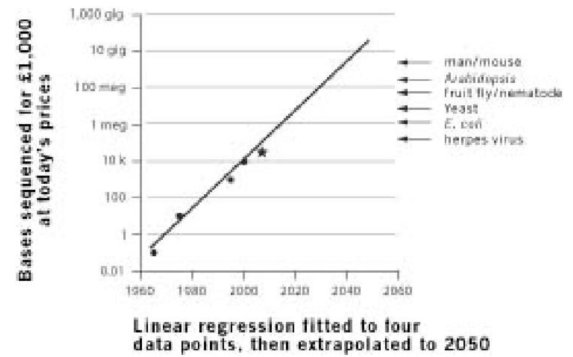
«Закон Ходгкина»
Наиболее масштабная попытка в этом направлении до настоящего времени была сделана группой, связанной с Дэвидом Хиллисом, братом Дэнни Хиллиса, который был основателем одного из первых суперкомпьютеров. Схема Хиллиса делает диаграмму дерева более компактной, оборачивая ее по кругу. Вы не увидите промежутка, где два конца почти встречаются, но он находится между "бактериями" и "археями". Чтобы понять, как работает эта круговая диаграмма, посмотрите на очень упрощенную версию, татуированную на спине доктора Клэр Д"алберто из университета Мельбурна, зоологический энтузиазм которой далеко не поверхностен. Клэр любезно позволила мне воспроизвести фотографию в этой книге (см. цветную страницу 25). Ее татуировка включает маленькую выборку из восьмидесяти шести видов (число конечных ветвей). Вы можете видеть промежуток в круглой диаграмме, и представить себе круг развернутым. Меньшее число рисунков вокруг края стратегически выбрано из бактерий, протозоев, растений, грибов, и четырех типов животных. Позвоночные животные представлены справа лиственным морским драконом, удивительной рыбой, защищаемой своим сходством с морскими водорослями. Круглая диаграмма Хиллиса такая же, за исключением того, что в ней три тысячи видов. Их названия, представленные вокруг внешнего края круга выше, слишком маленькие, чтобы прочесть - хотя Homo sapiens услужливо отмечен "Вы здесь". Вы можете составить представление, насколько разрежена выборка дерева даже в этой огромной диаграмме, если я скажу Вам, что самые близкие родственники людей, которые могли быть вписаны в круг, это крысы и мыши. Млекопитающие должны бы были быть существенно прорежены, чтобы соответствовать степени охвата во всех других ветвях дерева. Только представьте себе попытки начертить подобное дерево с десятью миллионами видов вместо этих трех тысяч, охваченных здесь. И десять миллионов - не самая радикальная оценка количества существующих видов. Стоит скачать дерево Хиллиса с его вебсайта (см. сноски), а затем напечатать его в виде настенного плаката на листе бумаги, который, как они рекомендуют, должен быть по крайней мере 54 дюйма шириной (хорошо бы еще больше).
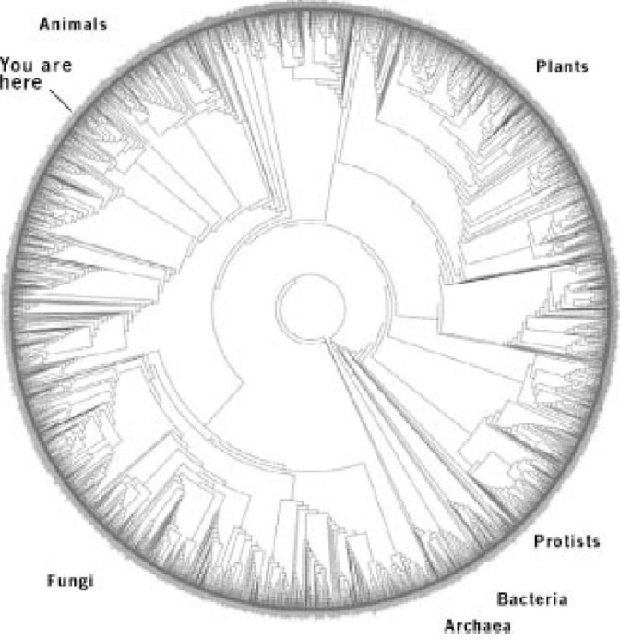
График Хиллиса
МОЛЕКУЛЯРНЫЕ ЧАСЫ
Теперь, когда мы заговорили о молекулах, у нас осталось незавершенное дело из главы про эволюционные часы. Там мы рассмотрели годичные кольца и различные виды радиоактивных часов, но мы отсрочили рассмотрение так называемых молекулярных часов, пока не узнаем о некоторых других аспектах молекулярной генетики. Время пришло. Думайте об этом разделе как о приложении к главе про часы.
Молекулярные часы исходят из того, что эволюция истинна, и что она продолжается с довольно постоянной скоростью в течение геологического времени, чтобы использоваться сама по себе как часы, при условии, что она может быть откалибрована, используя окаменелости, которые в свою очередь калиброваны радиоактивными часами. Так же, как свечные часы предполагают, что свечи сгорают с постоянной и известной скоростью, а водяные часы предполагают, что вода вытекает из ведра со скоростью, которая может быть откалибрована, а высокие напольные часы с маятником предполагают, что маятник качается с постоянной скоростью, так же молекулярные часы предполагают, что существуют определенные аспекты самой эволюции, которые происходят с постоянной скоростью. Эта постоянная скорость может быть откалибрована по тем частям эволюционной летописи, которая хорошо задокументирована (доступными датированию по радиоактивности) ископаемыми. После калибровки молекулярные часы могут использоваться для других частей эволюции, которые не задокументированы ископаемыми. Например, они могут использоваться для животных, которые не имеют твердых скелетов и редко фоссилизируются.
Хорошая идея, но что дает нам право надеяться, что мы найдем эволюционные процессы, которые бы шли с постоянной скоростью? Действительно, многие свидетельства предполагают, что скорости эволюции очень различаются. Задолго до современной эры молекулярной биологии Дж.Б.С.Холдейн предложил "дарвин" в качестве меры эволюционной скорости. Предположим, что за эволюционное время некоторая измеряемая черта животного изменяется в устойчивом направлении. Например, предположим, что средняя длина ног увеличивается. Если в течение миллиона лет длина ног увеличивается в e раз (2.718..., число, выбранное по причинам математического удобства, во что нам вникать не нужно), то говорят, что скорость эволюционного изменения равна одному дарвину. Сам Холдейн оценил скорость эволюции лошади приблизительно в 40 милидарвинов, в то время как эволюция домашних животных при искусственном отборе предположительно измеряется килодарвинами. Скорость эволюции гуппи, пересаженных в ручей без хищников, как описано в Главе 5, была оценена в 45 килодарвинов. Эволюция "живых ископаемых", таких как Lingula (страница 140), должна, вероятно, измеряться в микродарвинах. Вы поняли: скорости эволюции того, что можно увидеть и измерить, вроде ног и клювов, весьма изменчивы.
Если скорости эволюции настолько изменчивы, каким образом мы можем надеяться использовать их в качестве часов? Здесь приходит на помощь молекулярная генетика. На первый взгляд не ясно, как такое может быть. Когда измеряемые черты, такие как длина ноги, эволюционируют, то, что мы видим, является внешним и явным проявлением лежащего в их основе генетического изменения. Как тогда может быть, что скорости изменений на молекулярном уровне обеспечивают хорошие часы, в то время как скорость эволюции ноги или крыла -нет? Если ноги и клювы подвергаются изменению со скоростью в пределах от микродарвинов до килодарвинов, почему молекулы должны быть более надежными в качестве часов? Ответ в том, что генетические изменения, проявляющиеся во внешней и видимой эволюции - такие как ноги и руки - являются очень маленькой верхушкой айсберга, и они - верхушка, подверженная существенному влиянию переменчивого естественного отбора. Большинство генетических изменений на молекулярном уровне нейтрально и, можно поэтому ожидать, происходит со скоростью, не зависящей от полезности, и может даже быть приблизительно постоянной в пределах отдельно взятого гена. Нейтральное генетическое изменение не оказывает никакого воздействия на выживание животного, а это - полезная верительная грамота для часов. Ведь гены, которые влияют на выживание, положительно или отрицательно, как ожидается, эволюционируют с измененной скоростью, отражающей это.
Когда нейтральная теория молекулярной эволюции была впервые предложена, среди прочих, великим японским генетиком Моту Кимурой, она была спорной. Один ее вариант теперь получил широкое признание и, не вдаваясь здесь в подробные доказательства, я собираюсь принять его в этой книге. Поскольку я имею репутацию архи-"адаптациониста" (предположительно помешанного на естественном отборе как главной или даже единственной движущей силой эволюции), Вы можете быть уверены, что, если даже я поддерживаю нейтральную теорию, маловероятно, что многие другие биологи выступят против нее!
Нейтральная мутация - мутация, которая, хотя и легко измерима молекулярно-генетическими методами, не подлежит естественному отбору, ни положительному, ни отрицательному. "Псевдогены" нейтральны по одной причине. Они являются генами, которые когда-то делали что-то полезное, но теперь отодвинуты в сторону и никогда не транскрибируются или не транслируются. Для благополучия животного их с таким же успехом могло бы не существовать вообще. Но с точки зрения ученых, они очень даже существуют, и они -именно то, что нам нужно для эволюционных часов. Псевдогены -только один класс тех генов, которые никогда не транслируются в эмбриологии. Существуют и другие классы, которым ученые отдают предпочтение для молекулярных часов, но я не буду вдаваться в подробности. Для чего полезны псевдогены - так это чтобы ставить в неловкое положение креационистов. Они испытывают даже их креативную изобретательность, чтобы придумать убедительную причину, почему разумный проектировщик должен был создать псевдоген - ген, который абсолютно ничего не делает и всячески создает видимость того, что он - устаревшая версия гена, который некогда что-то делал - разве что, если только он нарочно не задался целью нас одурачить.
Оставляя в стороне псевдогены, существует замечательный факт, что с тем же успехом можно было бы обойтись без большей части генома (95 процентов в случае людей). Нейтральная теория применима даже ко многим из генов из оставшихся 5 процентов - генам, которые читаются и используются. Она применима даже к генам, жизненно важным для выживания. Здесь я должен выразиться яснее. Мы не говорим, что ген, к которому применима нейтральная теория, не оказывает никакого эффекта на тело. Мы лишь говорим, что мутантная версия гена имеет точно такой же эффект, как и немутировавшая версия. Независимо от того, насколько важным или неважным будет сам ген, мутировавшая версия имеет такой же эффект, как немутировавшая версия. В отличие от псевдогенов, где сам ген может должным образом быть описан как нейтральный, мы сейчас говорим о случаях, когда только мутации (то есть изменения в генах) могут строго быть описаны как нейтральные, но не сами гены.
Мутации могут быть нейтральными по различным причинам. Код ДНК - "вырожденный код". Это технический термин, означающий, что некоторые кодовые "слова" являются точными синонимами друг друга. Когда ген мутирует в один из его синонимов, Вы можете с тем же успехом не утруждаться называть это мутацией вообще. Действительно, это не мутация, насколько это касается последствий для тела. И по той же причине это не мутация вообще, насколько это касается естественного отбора. Но это является мутацией, насколько это касается молекулярных генетиков, так как они могут увидеть ее, используя свои методы. Это как если бы я должен был изменить шрифт, в котором я пишу слово, скажем, кенгуру на кенгуру. Вы все еще можете прочитать слово, и оно все еще означает то же самое австралийское прыгающее животное. Изменение шрифта с Minion на Helvetica обнаружимо, но не связано со значением.
Не все нейтральные мутации столь же нейтральны как эта. Иногда новый ген транслирует другой белок, но "активный центр" (вспомните точные формы "впадин", которые мы встретили в Главе 8) нового белка остается тем же, что и у старого. Следовательно, нет буквально никакого эффекта на эмбриональное развитие тела.
Немутировавшая и мутировавшая форма гена все еще являются синонимами, насколько это касается их эффектов на тела. Также возможно (хотя такие "ультрадарвинисты" как я склонны противиться этой идее), что некоторые мутации действительно изменяют тело, но таким образом, что это так или иначе не оказывает никакого эффекта на выживание.
Итак, подводя итог нейтральной теории, сказать, что ген или мутация "нейтральны", не обязательно означает, что сам ген бесполезен. Он может быть жизненно важным для выживания животного. Это лишь означает, что мутантная форма гена, который может быть, а может и не быть важным для выживания, не отличается от исходной формы по производимым ею эффектам (которые могут быть очень важными) на выживание. Ситуация такова, что, вероятно, не будет неправдой сказать, что большинство мутаций нейтральны. Они не обнаружимы естественным отбором, но обнаружимы молекулярными генетиками; и это - идеальное сочетание для эволюционных часов.
Ничто из этого не должно принижать крайне важной верхушки айсберга - меньшинство мутаций, которые не нейтральны. Именно они отбираются, положительно или отрицательно, в эволюции усовершенствования. Это те мутации, эффекты которых мы фактически видим - и естественный отбор их "видит" тоже. Это те мутации, отбор которых придает живым существам их поразительную иллюзию дизайна. Но, когда мы говорим о молекулярных часах, нас интересует остальная часть айсберга -нейтральные мутации, которых большинство.
С течением геологического времени геном подвергается ливню износа в форме мутаций. В той небольшой части генома, где мутации действительно имеют значение для выживания, естественный отбор скоро избавляется от плохих и благоприятствует хорошим. С другой стороны, нейтральные мутации просто накапливаются, безнаказанно и незаметно для всех, кроме молекулярных генетиков. И теперь мы нуждаемся в новом техническом термине: закрепление [или фиксация]. У новой мутации, если она по-настоящему новая, будет низкая частота в генофонде. Если вы вновь вернетесь к генофонду миллион лет спустя, возможно, что мутация увеличится в частоте до 100 процентов или близко к этому. Если это происходит, мутация, как говорят, "закрепилась". Мы больше не будем думать ней как о мутации. Она стала нормой. Очевидный путь для мутации, чтобы закрепиться - чтобы естественный отбор ей благоприятствовал. Но есть и другой способ. Она может закрепиться случайно. Так же, как, как благородная фамилия может однажды угаснуть из-за отсутствия наследников, так же и альтернативные варианты той мутации, о которой мы говорим, могут просто случайно исчезнуть из генофонда. Сама мутация может стать частой в генофонде благодаря той же самой удаче, которая заставила Смит стать самой распространенной фамилией в Англии. Конечно, намного более интересно, если ген закрепляется по серьезной причине - а именно благодаря естественному отбору - но это может также произойти и случайно, на достаточно большом числе поколений. И геологического времени вполне достаточно для нейтральных мутаций, чтобы закрепляться с предсказуемой скоростью. Скорость, с которой они это делают, варьируется, но она характерна для конкретных генов, и при условии, что большинство мутаций нейтрально, это именно то, что делает возможными молекулярные часы.
Закрепление имеет значение для молекулярных часов, потому что "закрепившиеся" гены - это гены, на которые мы смотрим, когда сравниваем двух современных животных, чтобы попытаться оценить, когда произошел раскол между их предками. Закрепившиеся гены -это гены, характеризующие вид. Это гены, которые едва ли не универсальны в генофонде. И мы можем сравнить гены, закрепившиеся у одного вида, с генами, закрепившимися у другого, чтобы оценить, насколько недавно разделились два вида. Есть осложнения, в которые я не буду вдаваться, потому что мы с Иеном Вонгом в полной мере обсуждали их в эпилоге к "Рассказу Бархатного Червя". С оговорками и с различными важными корректирующими факторами, молекулярные часы работают.
Так же, как радиоактивные часы тикают с весьма различными скоростями, с периодами полураспада в пределах от долей секунды до десятков миллиардов лет, так же различные гены обеспечивают удивительный диапазон молекулярных часов, подходящих для определения времени эволюционных изменений в масштабах от миллиона до миллиарда лет и всех промежуточных периодов. Так же, как у каждого радиоактивного изотопа есть свой характерный период полураспада, так же у каждого гена есть характерная скорость обновления - скорость, с которой новые мутации обычно фиксируются благодаря случайности. Для генов гистонов характерно обновление со скоростью одной мутации в миллиард лет. Для генов фибринопептида в тысячу раз быстрее, со скоростью обновления в одну новую мутацию, закрепляющуюся за миллион лет. Цитохром-C и набор генов гемоглобина имеют промежуточные скорости обновления, со временем фиксации, измеряемой в пределах от миллионов до десятков миллионов лет.
Ни радиоактивные, ни молекулярные часы не тикают размеренно, как маятниковые или ручные часы. Если бы Вы могли слышать их тиканье, они бы звучали как счетчик Гейгера, собственно для радиоактивных часов счетчик Гейгера - именно то, что Вы бы использовали, чтобы их услышать. Счетчик Гейгера не тикает размеренно, как часы; он тикает беспорядочно, странными, запинающимися порывами. Так звучали бы мутации и фиксации, если мы могли бы услышать их в очень большом геологическом масштабе времени. Но независимо от того, запинается ли он как счетчик Гейгера или тикает метрономом как часы, важная особенность хранителя времени в том, что он должен тикать с известной средней скоростью. Так действуют радиоактивные часы, и так действуют молекулярные часы. Я представил молекулярные часы, сказав, что они исходят из факта эволюции и поэтому не могут использоваться в качестве ее доказательства. Но теперь, поняв, как работают эти часы, мы можем видеть, что я был слишком пессимистичным. Само существование псевдогенов - бесполезных, нетранскрибируемых генов, которые имеют заметное сходство с используемыми генами - является прекрасным примером способа, которым животные и растения носят в себе свою историю, написанную повсюду в них. Но это - тема, которая должна подождать до следующей главы.

